Schlüsselbegriffe: Viscum album L., Loranthus europaeus L., Wirtspflanzen, Stadtbäume, Verteilungsmodelle, Wien, Graz
Available at https://doi.org/10.53203/fs.2501.2
See below the issue 1/2025 as E-Paper or have a look at our E-Paper archive dating back to 1955.
Abstract
Mistletoe species are part of the biodiversity of cities, forests and other tree-covered areas, but as a hemiparasite they are also a possible threat to trees, in particular in view of increasing drought stress as a consequence of climate change. The distribution of mistletoe largely depends on the woody host species and their growing density. The tree cadasters of the Austrian cities Vienna and Graz were analysed, as well as the data of the Austrian National Forest Inventory around Vienna for the inventory cycles 2000-2002, 2007-2009 and 2016-2021. Fieldwork and visual assessments of the trees were conducted to determine the prevalence of mistletoe infection. The most common host trees for mistletoe in Vienna and Graz (Acer, Tilia, Populus) and Austrian forests (Pinus, Quercus) were identified and the distribution of the various mistletoe species is described. The factors determining the presence of mistletoes within urban areas and surrounding forests were analysed. Our results allow to predict the spread of particular mistletoe species to adjacent areas. Our study also identified mistletoe-resistant trees, that remain healthy and live longer and may help mitigate the effects of climate change.
Zusammenfassung
Mistelarten sind Teil der biologischen Vielfalt in Städten, Wäldern und anderen baumbestockten Flächen, stellen als Hemiparasiten aber auch eine mögliche Bedrohung für Bäume dar, insbesondere im Hinblick auf zunehmenden Trockenstress als Folge des Klimawandels. Die Verbreitung der Mistel hängt weitgehend von der Gehölzart und dem Vorkommen dieser Wirtsarten ab. Ziel dieser Studie war es, Mistelarten in städtischen Gebieten und Wäldern zu identifizieren. Dafür wurden die Baumkataster von Wien und Graz sowie die Daten der Österreichischen Waldinventur (ÖWI) um Wien für die Perioden 2000–2002, 2007–2009 und 2016–2021 analysiert. Es wurden auch Feldarbeiten zur Erfassung von befallenen Bäumen durchgeführt, um die Intensität des Mistelbefalls zu ermitteln. Als Ergebnis der Arbeit wurden die häufigsten Wirtsbaumarten der Mistel in Wien und Graz (Acer, Tilia, Populus) und Österreichs Wäldern (Pinus, Quercus) identifiziert und die Besonderheiten der Verbreitung der verschiedenen Mistelarten aufgezeigt. Die Einflussfaktoren der Ausbreitung der Mistel in städtischen Gebieten und umgebenden Wäldern wurden nachgezeichnet. Anhand der gewonnenen Daten ist es möglich, die Ausbreitung bestimmter Mistelarten in angrenzende Gebiete vorherzusagen. Die Ergebnisse dieser Arbeit ermöglichen auch, mistelresistente Bäume auszuwählen, die durch Vitalität und Langlebigkeit einen Beitrag zur Milderung der Auswirkungen des Klimawandels leisten können.
1 Introduction
Mistletoes are an integral part of the biodiversity and the flora of Europe (Grazi and Urech, 1985; Zuber, 2004). The diverse interest in mistletoes is reflected in numerous studies (e.g. Krasylenko et al., 2019; Valle et al., 2021; Walas et al., 2022). The forest and urban green area infected by mistletoe in Europe has been increasing in recent years (Iszkuło et al., 2020; Thomas et al., 2022), and the prevalence of infections is expected to increase significantly (Dobbertin et al., 2005).
In Austria, two mistletoe species are frequently found, Viscum album L. and Loranthus europaeus L. Viscum album, which is presented by three subspecies: V. album subsp. album L., V. album subsp. abietis (Wiesb.) Abrom., V. album subsp. austriacum (Wiesb.) Vollm. All subspecies of V. album are evergreen and carry white fruits, while L. europaeus is deciduous and bears yellow berries (Glatzel et al., 2016).
All mistletoe species are important in natural ecosystems, but are hemiparasites. Mistletoes obtain water, nitrogen, nutrients and a significant portion of carbohydrates from their hosts. Scientific research by many authors describes the diverse negative effects on the physiological processes and growth and development of woody plants by mistletoes (Idžojtić et al., 2008; Türe et al., 2010; Kleszken et al., 2022).
Mistletoe can cause severe drought stress to trees in dry and hot seasons, thereby playing a potent role in increasing their mortality rate (Ančić et al., 2014; Mutlu et al., 2016). Various mistletoe species can lead to a reduction of leaf growth, vitality and longevity, to yield loss, and to the loss of decorative effect of the host trees (Catal and Carus, 2011; Barbu, 2012). Mistletoe infection also enhanced tree canopy defoliation (Dobbertin et al., 2005). The productivity of affected forest stands can decrease, as infected pine trees were reported to have lower increment and radial growth compared to non-infected trees (Bilgili et al., 2020). V. album had also a negative impact on the number and size of cones and seed quality of Pinus sylvestris L. (Pilichowski, 2018). Severely infeсted trees responded stronger to drought stress than non-infeсted trees (Sanguesa-Barreda, 2013), because mistletoe leaves showed high transpiration rates, even when the host is under severe drought stress (Aukema, 2003; Escher, 2008) and mistletoes maintain a higher relative water content than their host (Strong and Bannister, 2002).
Trees generally perform a wide range of ecosystem functions, which are compromised under poor health condition. Several factors affect tree health in today's environment of climate change. Selecting trees resistant to various negative factors, including to pests such as mistletoe, has thus gained importance (Alvarado-Rosales and Saavedra-Romero, 2021). It is key not only to know the species of mistletoe and host trees, but also to know the possible trajectories of their further spread. Deeper knowledge about the features of tree infection with mistletoes will allow us to counteract the uncontrolled spread of this hemiparasite. Compared to other tree pests, mistletoes develop and spread relatively slowly (Hawksworth et al., 1991). There is a lack of research in Europe into driving factors and distribution of mistletoe spread, allthough the increase in mistletoe occurrence and abundance is evident in many European countries. The level of mistletoe infection of trees in forests is predicted to increase (Kollas et al., 2018).
Establishing a better understanding of the dynamics of mistletoe abundance is necessary for developing conservation and management strategies for mistletoe-host complexes (Kubíček et al., 2018). The aim of this study was to determine and compare the abundance of mistletoes on host tree species, the infection rate, and to identify factors affecting the distribution of mistletoe species in urban and forest conditions. The results of the study are useful for the prediction of future spread of mistletoes, for modeling the infection of host species and the migration of certain mistletoe species from the city to the adjacent forest areas, and vice versa.
2 Materials and Methods
This research is based on the analysis of tree cadastres of the Cities of Vienna and Graz (Austria), Austrian National Forest Inventory (NFI) data (BFW, 2022) as well as on specific field assessments. The data from Vienna was provided by the Wiener Stadtgärten in November 2022, the data from Graz by Graz Holding in January 2023. The NFI data was provided by the Department of Forest Inventory of the Austrian Research Centre for Forests (BFW) in March 2023. The Austrian NFI conducts comprehensive assessments at regular time intervals on a systematic grid of permanent sample plots (Gabler and Schadauer, 2008; Gschwantner et al., 2010, 2016). The sample plots are arranged at the corners of square-shaped clusters with a side-length of 200 m. The sample plots consist of a small circular plot of 21.2 m², an angle count sample (Bitterlich, 1948), and a large circular plot of 300 m². The latter is the basis for the assessment and attribution of stand- and site-specific variables that include, among others, forest management type, growth classes, age classes, sea level, as well as the occurrence of tree and shrub species, and the presence/absence of mistletoes. The species occurrence assessments distinguish four vegetation layers, an upper tree layer 1, lower tree layer 2, shrub layer and ground vegetation layer (Hauk et al. 2016). For the present study, the NFI sample plot and mistletoe occurrence data for the cycles 2000-2002, 2007-2009 and 2016-2021 within a 50 km–circle around Vienna were used.
The shapefiles of the city boundaries for Figure 1 were obtained from https://www.data.gv.at/. The maps (Figure 5) were created using the R (2023) software.
To evaluate mistletoe infection prevalence, the fraction of trees infected by V. album and L. europaeus per species (or in total) was used. It was determined using the following formula (Lech et al., 2020):
F = 100 * n/N, (1)
F - frequency of V. album / L. europaeus, %; n - number of trees infected by V. album / L. europaeus among trees of a given species (or of all species together); N - total number of trees of a given species (or of all species together).
To determine if there is a statistically significant difference in the mistletoe distribution in the forest, we applied the Z-test for proportions (Zar, 2010). Pairwise comparisons of the mistletoe distribution percentages were performed across three periods (2000-2002, 2007-2009, and 2016-2021).
There are several ways described for assessing the infection of urban greening and forests in scientific literature. Different authors, based on their specific purposes, have suggested various classifications of infection prevalence (Kolodziejek et al., 2013; Bilonozhko et al., 2022). Based on the analysis of the NFI data, we developed our own scale of mistletoe infection prevalence. It comprises low (up to 1 %), medium (1 - 4.0 %), high (4.1 - 6.0 %) and very high (more than 6.1 %) infection rates.
The data was analysed with Excel statistical functions using with histograms to visualize the distribution of data based on Microsoft Office XP software, and Statistical Package for Social Sciences, version 11.5 (SPSS Inc., Chicago, 2002).
3 Results

Table 1: The most common and the most frequently infected woody species in Vienna and Graz.
Tabelle 1: Die häufigsten und am häufigsten infizierten Gehölzarten in Wien und Graz.
A more comprehensive analysis for Vienna is given in a companion manuscript (Bilonozhko et al., 2025). We concentrate here on reporting comparisons between the two cities and the situation in forests, and only give a brief overview for Vienna.
3.1 Mistletoe infection in Vienna
There are 213,841 trees recorded in the tree cadastre of Vienna. The overall mistletoe infection frequency rate is 3.05 % (6,537 individuals). Two different species of mistletoe were found in Vienna: V. album subsp. album (97 %) and L. europaeus (3 %). The diversity of woody species in Vienna is very rich and comprises 546 taxa, among which the most common are Acer campestre, A. platanoides, A. pseudoplatanus, Aesculus hippocastanum, Celtis australis, Fraxinus excelsior, Pinus nigra, Platanus x acerifolia, Tilia cordata and T. platyphyllos (Table 1). A significant number of species (114) were infected by V. album.
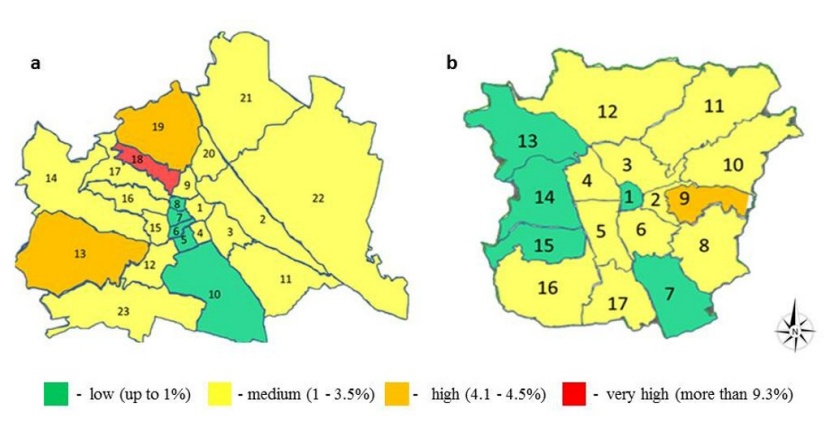
Figure 1: Schemes of the districts in Vienna (a) and Graz (b). The colouring indicates the level of mistletoe infection of urban trees. The increase in the number of infected plants is shown from green to red.
Abbildung 1: Karte der Bezirke in Wien (a) und Graz (b). Die Farben zeigen den Grad des Mistelbefalls in städtischen Bäumen an. Die Zunahme der Zahl der befallenen Pflanzen ist von grün nach rot dargestellt.
In Vienna, the most intensively infected species by V. album is Juglans nigra, where 35.2 % of the 193 trees are infected. Populus balsamifera also has a high degree of infection (26.3 %), although only 57 individuals are growing in Vienna. There are 1,014 trees of Celtis occidentalis planted, 21.5 % of them are affected. These examples demonstrate the selectivity of mistletoes with regard to host tree species. Seen from another angle, the occurrence of mistletoes also depends highly on the presence of suitable host tree species.
Species with a high degree of infection are key in the spread of mistletoes. Despite a relatively high presence of the pest in the cities, not all tree species are susceptible to mistletoe. There are deciduous species well represented by a significant number of trees, that appear to be resistant to V. album subsp. album: Ailanthus altissima, Catalpa bignonioides, Corylus colurna, Fagus sylvatica, Gleditsia triacanthos, Koelreuteria paniculata, Liriodendron tulipifera, Magnolia sp., Morus alba, Paulownia tomentosa, Platanus orientalis, Platanus x acerifolia, Sambucus nigra and Sophora japonica. Also, coniferous species appear resistant to V. album subsp. album in Viennese urban trees.
Host species of L. europaeus are only found in the genus Quercus, in particular Q. cerris, Q. macrocarpa, Q. petraea, Q. robur and Q. rubra. L. europaeus can infect up to 86 % of trees in clustered (group) plantings such as parks and squares.
In the various districts of Vienna, the number of V. album and L. europaeus and the dynamics of their populations differ significantly. This usually depends on the total area of the district, the number of trees present, their growth density, and the species composition of the trees. The average level of tree infection by mistletoe in Vienna is 3.05 %, but there are districts in Vienna (5, 6, 7, 8 and 10) where less than 1 % of the trees are infected (Fig. 1). Districts 13, 18, 19 are infected above the average level (these numbers do not include some large parks not covered by the cadaster data; but heavy infections there would likely spread in the surroundings; see further below). All other districts have a percentage of infections below average for Vienna. The presence of the main mistletoe host tree species in significant numbers in certain districts causes a higher overall infection percentage.
3.2 Mistletoe infection in Graz
There are 18,475 trees recorded in the tree cadaster of Graz. The number of trees affected by mistletoe is 285 (1.5 %). Woody species diversity is represented by 223 taxa, among them the most common species are Acer platanoides, A. pseudoplatanus, Aesculus hippocastanum, Carpinus betulus, Fraxinus excelsior, Platanus x acerifolia, Populus nigra, Prunus serrulata, Sophora japonica and Tilia cordata. There are 36 species infected by V. album in Graz. The highest infection was found on Populus nigra, where 19.1 % of the 47 trees are infected. Malus floribunda also has a high degree of infection (18.4 %), although only 38 individuals are growing in Graz. There are 65 trees of Acer saccharinum planted, 15.4 % of them are affected. (Table 1). These are typical host trees in Graz and the main links in the chain of mistletoe spread to other species. Species that are not susceptible to mistletoe infection are the same as those in Vienna, but their percentage share in the species composition of Graz is significantly larger, which significantly reduces the overall level of infection in the city.
L. europaeus is absent in the urban environment of Graz, although several Quercus species are present there. The level of urban tree infection in Graz varies among districts, as already seen in Vienna (Figure 1). In districts 1, 7, 13, 14, and 15, the percentage of trees with mistletoe is very low. Districts 2, 3, 4, 5, 6, 8, 10, 11, 12, 16, and 17 show an average percentage of infection, while mistletoe is relatively widespread in the district 9 (4.2 %). Typically, the prevalence of infection depends on the density of tree presence and on the predominant species.
There is no significant mistletoe ‘problem’ in Graz, as the percentage of infected trees is low, which means there is no apparent threat to trees in general. At the same time, a good balance of biodiversity in the city is maintained, i.e., mistletoes are playing their role in supporting other species like birds. There is no tendency for a sharp increase. The local climate in Graz is somewhat different from Vienna (see below), due to the higher altitude of Graz.

Figure 2: Occurrence of mistletoe species on the NFI plots in the cycles 2000-2002, 2007-2009 and 2016-2021 (total number of plots 987).
Abbildung 2: Vorkommen von Mistelarten auf den NFI-Probeflächen in den Perioden 2000-2002, 2007-2009 and 2016-2021 (Anzahl der Plots mit Misteln in verschiedenen Erhebungsperioden, Gesamtzahl der Plots 987).
3.3 Mistletoe in Austrian forests
The forests around Vienna are dominated by a range of different tree species. In the north and north-west Vienna, the majority of forests are broadleaf-deciduous with Fagus, Quercus and Carpinus being dominant, and to a lesser extent, Fraxinus, Acer and Tilia. In the south and south-east, evergreen-coniferous forests, mostly Pinus, are more frequent. According to the Austrian NFI, mistletoe infection in most areas around Vienna is below 5 %, very rarely up to 10 %. Infections of trees in the forests are caused mainly by V. album (75 %) and to a lesser degree by L. europaeus (25 %).
Pinus sylvestris and P. nigra dominate the coniferous forests around Vienna and Viscum album subsp. austriacum is the most common hemiparasite in this area. V. album subsp. album is less common but is found on a variety of hosts: Acer campestre, A. negundo, A. pseudoplatanus, Alnus incana, Betula pendula, Carpinus betulus, Populus x canadensis, P. nigra, Robinia pseudoacacia, Salix sp. and Tilia platyphyllos. Occasionally V. album subsp. abietis occurs on Abies alba. Fagus sylvatica, very common in the Viennese woods along the edge of the cities are not susceptible to mistletoe and act as a barrier.
The most common host for L. europaeus, commonly called the oak mistletoe, around Vienna is Quercus petraea. Other oak species like Q. cerris, Q. pubescens, Q. robur and Q. rubra are less common and less affected.
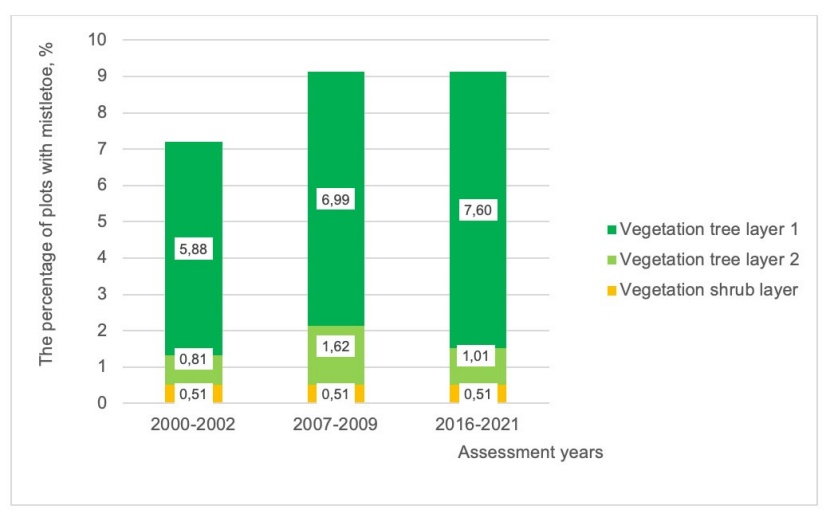
Figure 3: Distribution of mistletoe occurrences in the different vegetation layers of the NFI plots during the cycles 2000-2002, 2007-2009 and 2016-2021.
Abbildung 3: Verteilung des Mistelvorkommens in den verschiedenen Vegetationsschichten auf den NFI-Probeflächen in den Perioden 2000-2002, 2007-2009 und 2016-2021 (grün: Baumschicht 1, hellgrün: Baumschicht 2, orange: Strauchschicht).
The number of NFI plots with mistletoe occurrence has increased between the NFI cycles 2000-2002 and 2016-2021. In particular, infections by both species (V. album and L. europaeus) increase, which is due to the presence of a wide range of host species (Figure 2). The results of the pairwise comparisons of mistletoe distribution percentages showed that the differences between all comparisons were not statistically significant, indicating that the changes in mistletoe distribution across these periods are not substantial.
The age of mistletoe-infeсted stands is, in most cases, recorded as higher than 21 years. It is possible for mistletoe to establish in different vegetation layers, but mistletoe shows a preference for the upper tree layer (Figure 3).

Figure 4: Frequency of mistletoe infected trees by genera in Vienna and Graz.
Abbildung 4: Anteil der von Misteln befallenen Bäumen gegliedert nach Gattungen in Wien (blau) und Graz (gelb)
3.4 Mistletoe distribution forecast
The highest potential threat in the city of Vienna is posed by V. album subsp. album and L. europaeus. In Graz, V. album subsp. album is also the most common species. V. album subsp. austriacum is often found in forests around Vienna, V. album subsp. abietis is rare, since Abies sp. are not common urban trees.
The vast majority of trees with V. album subsp. album belong to the genera Acer, Betula, Malus, Populus, Robinia, and Tilia (Figure 4). Only 12.3 % of the infected trees in Vienna and 11.2 % in Graz belong to other genera. Most of the trees in Vienna are of the genus Acer, while Tilia trees are widely represented in Graz. Accordingly, tree species of these genera have the highest infection rate.

Figure 5: Occurrence of Viscum album (a) and Loranthus europaeus (b) in the Austrian forests according to the Austrian NFI 2016-2021 (probabilities from blue: low to red: high; black dots are inventory plots with mistletoe presence).
Abbildung 5: Vorkommen von Viscum album (a) und Loranthus europaeus (b) in den Österreichischen Wäldern nach der Österreichischen Waldinventur 2016–2021.
The cluster arrangement of main host species (for example Juglans nigra, Populus balsamifera and Celtis occidentalis in Vienna and Populus nigra in Graz) has the greatest impact on the prevalence of infection among these species.
Sustainable forest management also requires measures for forest protection, thus there is a need for monitoring and preventing the development of pests. The infection of forests with V. album in most areas of Austria has been determined to be insignificant (Figure 5). The areas with the highest degree of infection are located in the eastern part of Austria. Individual local patches of mistletoe infected trees do not currently have a significant impact on the stands, but they are sources of further spread for mistletoes.
L. europaeus only infects species of the genus Querсus, where the intensity can be quite high in some species of the genus. The hotspot for L. europaeus is north-eastern Austria, a rather flat and warm part of the country (Figure 5). In the rest of the country, oak stands in hills and mountains are unsuitable for L. europaeus. This mistletoe species seems to prefer a hotter and drier climate (see Discussion). Factors affecting mistletoe spread are mainly related to host species, their location, abiotic factors, and distribution vectors like birds.
Mistletoe distribution can be influenced in urban areas by planting more resistant tree species. In the forest, it is almost impossible to create conditions for stopping the spread of mistletoe due to high tree density, long rotation times, and the large scale of any operations.
4 Discussion
4.1 Climate change and reproduction of mistletoe
Climate change is having a significant impact on all living organisms, including on trees, as well as on the spread of mistletoe. Mistletoe abundance is increasing in all parts of their existing distribution ranges (Dobbertin and Rigling, 2006; Turner and Smith, 2016).
Forests are complex ecosystems, with many interacting ecosystem elements. In the absence of severe, significant stress factors, changes occur slowly, and the spread of mistletoe in the forest remains predictable.
The massive reproduction of mistletoes and increase in their abundance are likely caused by climate change, which has led to a significant decrease of the health status of woody plants and to the creation of more comfortable conditions for pests, including hemiparasitic plants (Walas et al., 2022; Jeffree and Jeffree, 1996). The number of mistletoes on trees increases with declining tree vitality (Baltazár et al., 2013; Bilonozhko et al., 2025). Cases of severe infection of forests have been described, such as fir forests with 67 % infection by V. album subsp. abietis (Tsopelas et al., 2004). Here it is interesting to note that the apparent cline in mistletoe infection from Northeastern Austria (heavy infections especially of L. europaeus) through Vienna to Graz (least infected) coincides with the climate – Northeastern Austria is the warmest and driest part, and the area around Graz receives more annual rainfall than Vienna and the Northeast (detailed climate data available from: https://www.zamg.ac.at/cms/de/klima/informationsportal-klimawandel/daten-download/copy_of_klimamittel; accessed 4 June 2024). Dobbertin et al. (2005) likewise discussed the possible upwards shift of mistletoes in forests due to climate warming.
4.2 Mistletoe distribution in cities
Trees in the urban environment often grow under stressful conditions, which make them more susceptible to pests, in particular to mistletoes (Díaz-Limón et al., 2016). Due to the wide diversity of woody species in cities, the number of host species infected by mistletoe can make up to 500 (Barney et al., 1998; Szmidla et al., 2019; Thomas et al., 2022). Moreover, some species of trees and shrubs are more intensively infected than others (Krasylenko et al., 2020). The most common host species of V. album subsp. album in European cities are Acer platanoides, A. saccharinum, Betula pendula, Robinia pseudoacacia, Salix alba, Tilia cordata, Populus nigra (Bilonozhko et al., 2022). Scientists also record new (previously undescribed) host species of mistletoes (Idžojtić et al., 2006). It is often noted that introduced and exotic tree species are more heavily infected than native ones (Kolodziejek et al., 2013; Silva and Fadini, 2017; for a detailed analysis of this aspect in Vienna, see Bilonozhko et al., 2025). Individual mistletoe species have a narrow range of host trees, such as L. europaeus (Zebec and Idžojtić, 2006). However, the principle by which some host plants are affected more abundantly than others has not been definitively clarified (Becker, 2000; Lech et al., 2020).
4.3 Mistletoe distribution in forests
As mistletoe is a heliophyte, mistletoe is often found on edges of the forests (Lorenc and Véle, 2022) and less prevalent on the trees deeper within the forest. Solitary trees have twice the infection load compared to trees located within closed forest canopies. Lower canopy density or proximity to open space is a very important factor for mistletoes. The prevalence of the hemiparasitic is increasing with woodland fragmentation (Barbosa et al., 2016). Polish researchers described main stand characteristics connected to mistletoe infection (Wójcik and Kędziora, 2020): Kraft class (Assmann, 1970), crown density, site index and age class. In Polish forests, it was found that the trees with the highest number of mistletoe bushes grew under the best light conditions. The most heavily infected trees were the tallest and oldest, that have higher diameter and higher defoliation (Lorenc and Véle, 2022), growing in low density, and having the biggest crowns.
4.4 Mistletoe as a part of the ecosystem
The specific impact of mistletoes on forest ecosystems is a gradual change of the stand. A decrease of tree vitality in forests leads to thinner crowns and shortens the host's life expectance, which in turn increases the availability of light and contributes to mistletoe spread (Mellado and Zamora, 2017; Dzyba, 2022). It was found that in infected forest stands, basal area increment can decrease up to 27 % during nine years (Kollas et al., 2018). The role of mistletoes in ecosystems is still controversial. On the one hand, they are considered a major biotic stressor for host trees, which can cause the death of trees and whole stands. On the other hand, mistletoes play an important ecological role in increasing biodiversity (Griebel et al., 2017; Krasylenko et al., 2020). Recently, the role of mistletoes has been described as an ecosystem engineer species (Mathiasen et al., 2008; Maul et al., 2019). Many mistletoes have closely associated species including birds, invertebrates and fungi. Some insects are fully dependent on mistletoe (Briem et al., 2016; Briggs, 2021). In some cases, mistletoe contributes to an increase in biodiversity by providing structural and nutritional resources (Watson and Herring, 2012). Mistletoes play a key role in intensifying soil resource availability, as they are regulating the abundance of soil microbial communities (Mellado et al., 2016). The experimental removal of mistletoes in the forest has led to quantitative ecosystem changes, highlighting the importance of the connections that determine the functioning of the ecosystem (Watson, 2015). A better understanding of mistletoe and host interaction can be used in forest management and for conservation of biodiversity, and of rare and endangered species (Glatzel and Geils, 2008).
The main seed dispersers of mistletoe are birds. In Europe, mistletoe is mainly consumed by thrushes (Turdus iliacus, T. merula, T. pilaris, T. philomelos, T. torquatus, T. viscivorus), tits (Cyanistes caeruleus, Parus major, Periparus ater), jay (Garrulus glandarius), blackcap (Sylvia atricapilla), robin (Erithacus rubecula) (Figarski, 2009; Varga et al., 2012; Thomas, 2022) and waxwing (Bombycilla garrulus) (Mellado and Zamora, 2014). Depending on the species, a bird can carry seeds up to 100 km. Mistletoe berries are an important source of food in winter and attractive to birds for a number of reasons: they remain on the plant for a long time and contain large amounts of carbohydrates. Birds also build nests in mistletoe bushes (Cooney, 2006). However, this is a very complex topic, as the foraging behaviour of various bird species can change with weather conditions (e.g. presence of snow); because of the migration behaviour of some birds in large swarms in winter (which may also change because of climate change); and because of the nutrition value of mistletoe berries for different species of birds (sometimes with a very fast passage through the intestinal tract) – all these aspects are analysed e.g. by Dieberger (1982).
There is a high probability that coniferous trees in cities that have not yet been infected with mistletoe are at a high risk. Birds can act as vectors, carrying berries with them. Vice versa, infection hotspots of V. album subsp. album in the city are a potential risk for the surrounding forests as a result of bird migration.
The distribution of mistletoes is influenced not only by climatic, but also by orographic factors. Altitude is the most significant factor of the distribution of mistletoes in mountains (Bilgili et al., 2020). Our research suggests that the limiting factor for the distribution of L. europaeus is altitude. We found that the number of species infected by mistletoe in Graz is significantly lower than in Vienna. L. europaeus is completely absent from Graz. Vienna is situated at 190 m, Graz at 353 m. In Graz, unlike in Vienna, mistletoes do not parasitize rare species. However, altitude and climate are interconnected, and it is difficult to discern these.
As mistletoes are long-lived, it can take decades before their harmful effects on the host and their spread become apparent. Therefore, it is necessary to regularly monitor mistletoe behaviour in space and time. The species composition of host trees in the city and outside the city is different. Monitoring should pay special attention to parks and green spaces in close proximity to forest areas.
5 Conclusions
Nowadays, a lot of scientific research focuses on biodiversity and trees resilient in climate change. The role of tree species in cities and forests as mistletoe hosts is underestimated and partially determines the future of the environment. The analysis of tree cadastres in Vienna and Graz, as well as the results of the forest inventory, allowed us to identify mistletoe species and their hosts in both urban and forest areas. There are many more tree species in urban green space than in forests. Therefore, mistletoe, as a tree pest, is widely present in urban trees. The main mistletoe host species in Austrian cities are Acer platanoides, A. saccharinum, Celtis occidentalis, Juglans nigra, Populus balsamifera, P. nigra, P. x canadensis, Robinia pseudoacacia, Tilia americana, T. cordata, Tilia x europaea. A significant concentration of V. album subsp. album in the city can contribute to its spread, further beyond the city limits into the surrounding forests.
The common mistletoe host species in forests are Pinus sylvestris and P. nigra. The coniferous mistletoe species (V. album subsp. austriacum) could inhabit urban trees in the future. L. europaeus will continue to spread in group and alley oak plantings. Due to birds, urban and forest trees can act as potential stepping stones for mistletoes in both directions.
By assessing the complexity of factors that can affect the occurrence and spread of mistletoe, it is possible to prevent or significantly slow down the rate of spread, and therefore reduce the affected area. We suggest considering tree species, which cannot be infected by mistletoe when choosing urban tree species for future planting. However, it is important to preserve mistletoes in safe quantities as an important part of biodiversity.
To monitor future population dynamics, it is important to keep precise records of mistletoe infections. It is also important to differentiate between the two species of mistletoe and to distinguish in statements and publications between various tree species and possibly even cultivars.
Conflicts of Interest
The authors declare no conflict of interest.
Author Contributions
Conceptualization, methodology was designed by Olha Tokarieva and Yuliia Bilonozhko; writing original draft preparation - Olha Tokarieva and Yuliia Bilonozhko; writing review and editing - Berthold Heinze and Thomas Gschwantner; project administration, funding acquisition, writing review and editing - Andrea Kodym. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This project (2022-0.710.109) was funded by the Federal Ministry of Agriculture, Forestry, Regions and Water Management via dafne.at. The BML supports applied, problem-oriented research in its area of expertise. Special thanks to Graz Holding and the Vienna City Gardens, especially to Mr. Robert Grill (Graz) and Mr. Clemens Weiss (Vienna) for the provision of the data and the professional cooperation. Many thanks also to Mr. Rainhard Hagen for the exchange of information. Heartfelt thanks to Alois Schuschnigg from BFW for supporting Ukrainian scientists.
References
Alvarado-Rosales D, Saavedra-Romero L (2021). Tree damage and mistletoe impact on urban green areas. Revista Árvore 45 (1): 4530. https://doi.org/10.1590/1806-908820210000030.
Ančić M, Pernar R, Bajić M, Seletković A, Kolić J (2014). Detecting mistletoe infestation on silver fir using hyperspectral images. iForest 7: 85−91.
Aukema JE (2003). Vectors, viscin, and Viscaceae: mistletoes as parasites, mutualists, and resources. Frontiers in Ecology and the Environment 1 (4): 212–219. https://doi.org/10.1890/1540-9295(2003)001[0212:VVAVMA]2.0.CO;2.
Assmann E (1970). The Principles of Forest Yield Study. Oxford. Pergamon Press.
Baltazár T, Pejchal M, Varga I (2013). Evaluation of European mistletoe (Viscum album L.) infection in the castle park in Lednice. Acta Universitatis Agriculturae et Silviculturae Mendelianae Brunensis 61 (6): 1565–1574. http://doi.org/10.11118/actaun201361061565.
Barbosa JM, Sebastián-González E, Asner GP, Knapp DE, Anderson C, Martin RE, Dirzo R (2016). Hemiparasite–host plant interactions in a fragmented landscape assessed via imaging spectroscopy and LiDAR. Ecological Applications 26: 55–66. https://doi.org/10.1890/14-2429.
Barbu CO (2012). Impact of white mistletoe (Viscum album ssp. abietis) infection on needles and crown morphology of silver fir (Abies alba Mill.). Notulae Botanicae Horti Agrobotanici Cluj-Napoca 40: 152–158. https://doi.org/10.15835/NBHA4027906.
Barney CW, Hawksworth FG, Geils BW (1998). Hosts of Viscum album. Eur. J. Forest Pathology 28: 187–208. https://doi.org/10.1111/j.1439-0329.1998.tb01249.x.
Becker H (2000). European mistletoe: Taxonomy, host trees, parts used, physiology. In: Büssing A (ed.). Mistletoe: The Genus Viscum. Hardwood Academic Publishers: Amsterdam, The Netherlands, pp 31–44. https://doi.org/10.1201/9780203304716.
BFW (2022). Österreichische Waldinventur. Waldinformationen aus erster Hand. Umfassend. Kompetent. Aktuell. www.waldinventur.at.
Bilgili E, Kadir CA, Baysal I (2020). The distribution of pine mistletoe (Viscum album ssp. austriacum) in Scots pine (Pinus sylvestris) forests: from stand to tree level. Scandinavian Journal of Forest Research 35 (1–2): 20–28. https://doi.org/10.1080/02827581.2020.1729402.
Bilonozhko YO, Kalafat LO Rabokon AM, Postovoitova AS, Privalikhin SM, Demkovych AE, Pirko YV (2022). Some characteristics of woody plants inhabited by Viscum album (Santalaceae). Ukrainian Botanical Journal 79 (6): 388–396 https://doi.org/10.15407/ukrbotj79.06.388.
Bilonozhko Y, Tokarieva O, Heinze B, Feichter J, Kodym A (2025). Mistletoe on urban trees in the city of Vienna, Austria. Urban Forestry & Urban Greening 128740. https://doi.org/10.1016/j.ufug.2025.128740.
Bitterlich W (1948). Die Winkelzählprobe. Allgemeine Forst- und Holzwirtschaftliche Zeitung 59: 4-5. (in German).
Briem F, Eben A, Gross J, Vogt H, (2016). An invader supported by a parasite: Mistletoe berries as a host for food and reproduction of Spotted Wing Drosophila in early spring. Journal of Pest Science 89 (3): 749–759. https://doi.org/10.1007/s10340-016-0739-6.
Briggs J (2021). Mistletoe, Viscum album (Santalaceae), in Britain and Ireland; a discussion and review of current status and trends. British & Irish Botany 3 (4): 419–454.
Catal Y, Carus S (2011). Effect of pine mistletoe on radial growth of Crimean pine (Pinus nigra) in Turkey. Journal of Environmental Biology 32: 263.
Cooney SJN, Watson DM, Young J (2006). Mistletoe nesting in Australian birds: a review. Emu – Austral Ornithology 106 (1): 1–12. https://doi.org/10.1071/MU04018.
Díaz-Limón MP, Cano-Santana Z, Queijeiro-Bolaños, ME (2016). Mistletoe infection in an urban forest in Mexico City. Urban Forestry & Urban Greening 17 (1): 126−134. https://doi.org/10.1016/j.ufug.2016.04.004.
Dieberger J 1982: Zoologische Komponente bei der Infektion und Verbreitung der Eichenmistel – vorläufige Ergebnisse. In: Mayer H (ed.), Der Eichenmistelbefall im Weinviertel. Veröff. Inst. Waldbau Univ. Bodenkultur Wien. (in German).
Dobbertin M, Hilker N, Rebetez M, Zimmermann N, Wohlgemuth T, Rigling A (2005). The upward shift in altitude of pine mistletoe (Viscum album ssp. austriacum) in Switzerland – the result of climate warming? International Journal of Biometeorology 50: 40–47.
Dobbertin M, Rigling A (2006). Pine mistletoe (Viscum album ssp. austriacum) contributes to Scots pine (Pinus sylvestris) mortality in the Rhone valley of Switzerland. Forest Pathology 36: 309–322.
Dzyba A (2022). Viscum album L. in protected areas of the Ukrainian Polissya. Forestry Ideas 1 (63): 254–267. (in Ukrainian).
Escher P, Peuke AD, Bannister P, Fink S, Hartung W, Jiang F, Rennenberg H (2008). Transpiration, CO2 assimilation, WUE, and stomatal aperture in leaves of Viscum album L: Effect of abscisic acid (ABA) in the xylem sap of its host. Plant physiology and biochemistry 46: 64–70.
Figarski T (2009). Selected aspects of wintering of Mistle Thrush Turdus viscivorus in the Kozienice Forest. Kulon 14: 1–7. (in Polish).
Gabler K, Schadauer K (2008). Methods of the Austrian Forest Inventory 2000/02 – Origins, approaches, design, sampling, data models, evaluation and calculation of the standard error. BFW-Berichte, p 121.
Glatzel G, Geils BW (2008). Mistletoe ecophysiology: host–parasite interactions. Botany 87 (1): 10–15.
Glatzel G, Richter H, Devkota MP, Amico G, Lee S, Lin R, Grabner M, Barlow BA, (2016). Foliar habit in mistletoe-host associations. Botany 95 (3): 219–229. https://doi.org/10.1139/cjb-2016-0161.
Grazi VG, Urech K (1985). Hyperparasitismus von Viscum album auf Loranthus europaeus als mögliches Bekämpfungsmittel gegen die Eichenmistelplage. Beitrage zur Biologie der Pflanzen 60: 467–474. (in German).
Griebel A, Watson D, Pendall E (2017). Mistletoe, friend and foe: synthesizing ecosystem implications of mistletoe infection. Environmental Research Letters 12. https://doi.org/10.1088/1748-9326/aa8fff.
Gschwantner T, Gabler K, Schadauer K, Weiss P (2010). National Forest Inventory Reports: Austria. In: National Forest Inventories: Pathways for Common Reporting. Springer Netherlands, pp 57–71.
Gschwantner T, Berger A, Büchsenmeister R, Hauk E (2016). National Forest Inventories Reports: Austria. In: C. Vidal, et al. (Eds.), National Forest Inventories – Assessment of wood availability and use. Springer International Publishing, Cham, pp 135-157.
Hauk E, Niese G, Schadauer K (2016). Instruktion für die Feldarbeit der Österreichischen Waldinventur 2016+ (04/2020). Bundesforschungszentrum für Wald (BFW). p 243. (in German). https://waldinventur.at/2016_Dienstanweisung_%C3%96WI_Fassung2020.pdf.
Hawksworth FG, Scharpf RF, Marosy M (1991). European mistletoe continues to spread in Sonoma County. California Agriculture 45: 39–40.
Idžojtić M, Kogelnik M, Franjić J, Škvorc Ž (2006). Hosts and distribution of Viscum album L. ssp. album in Croatia and Slovenia. Plant Biosystems 140: 50–55.
Idžojtić M, Pernar R, Glavaš M, Zebec M, Diminic D (2008). The incidence of mistletoe (Viscum album ssp. abietis) on silver fir (Abies alba) in Croatia. Biologia 63: 81–85. https://doi.org/10.2478/s11756-008-0014-2.
Iszkuło G, Armatys L, Dering M, Ksepko M, Tomaszewski D, Ważna A, Giertych MJ, (2020). Jemioła jako zagrożenie dla zdrowotności drzewostanów iglastych. Sylwan 164 (3): 226−236. https://doi.org/ 10.26202/sylwan.2019121.
Jeffree CE, Jeffree EP (1996). Redistribution of the potential geographical ranges of mistletoe and Colorado beetle in Europe in response to the temperature component of climate change. Functional Ecology 10: 562–577. https://doi.org/ 1010.2307/2390166.
Kleszken E, Purcarea C, Pallag A, Ranga F, Memete AR, Miere (Groza) F, Vicas SI (2022). Phytochemical profile and antioxidant capacity of Viscum album L. subsp. album and effects on its host trees. Plants 11: 3021. https://doi.org/10.3390/ plants11223021.
Kollas C, Gutsch M, Hommel R, Lasch-Born P, Suckow F (2018). Mistletoe-induced growth reductions at the forest stand scale. Tree Physiology 38 (5): 735–744. https://doi.org/10.1093/treephys/tpx150.
Kolodziejek J, Patykowski J, Kolodziejek R (2013). Distribution, frequency and host patterns of European mistletoe (Viscum album subsp. album) in the major city of Lodz. Biologia 68 (1): 55–64. https://doi.org/10.2478/s11756-012-0128-4.
Krasylenko Y, Sosnovsky Y, Atamas N, Popov G, Leonenko V, Janošíková K, Sytschak N, Rydlo K, Sytnyk D (2020). The European mistletoe (Viscum album L.): distribution, host range, biotic interactions, and management worldwide with special emphasis on Ukraine. Botany 98 (9): 499–516. https://doi.org/10.1139/cjb-2020-0037.
Krasylenko YA, Gleb RY, Volutsa OD (2019). Loranthus europaeus (Loranthaceae) in Ukraine: an overview of distribution patterns and hosts. Ukrainian Botanical Journal 76 (5): 406–417.
Kubíček J, Špinlerová Z, Michalko R, Vrška T, Matula R (2018). Temporal Dynamics and size effects of Mistletoe (Loranthus europaeus Jacq.) Infection in an Oak Forest. Austrian Journal of Forest Science 135 (2): 119–135.
Lech P, Żółciak A, Hildebrand R (2020). Occurrence of European mistletoe (Viscum album L.) on forest trees in Poland and its dynamics of spread in the period 2008–2018. Forests 11: 83. https://doi.org/10.3390/f11010083.
Lorenc F, Véle A (2022). Characteristics of Pinus sylvestris stands infected by Viscum album subsp. austriacum. Austrian Journal of Forest Science 139 (1): 31–50.
Mathiasen RL, Nickrent, DL, Shaw DC, Watson DM (2008). Mistletoes: pathology, systematics, ecology, and management. Plant Disease 92 (7): 988–1006. https:// doi.org/10.1094/PDIS-92-7-0988.
Maul K, Krug M, Nickrent DL, Müller KF, Quandt D, Wicke S (2019). Morphology, geographic distribution, and host preferences are poor predictors of phylogenetic relatedness in the mistletoe genus Viscum L. Molecular Phylogenetics and Evolution 131: 106–115.
Mellado A, Morillas L, Gallardo A, Zamora R (2016). Temporal dynamic of parasite-mediated linkages between the forest canopy and soil processes and the microbial community. New Phytologist 211: 1382–1392.
Mellado A, Zamora R (2014). Generalist birds govern the seed dispersal of a parasitic plant with strong recruitment constraints. Oecologia 176: 139–147. https://doi.org/10.1007/s00442-014-3013-8.
Mellado A, Zamora R (2017). Parasites structuring ecological communities: The mistletoe footprint in Mediterranean pine forests. Functional Ecology 31 (11): 2167–2176. https://doi.org/10.1111/1365−2435.12907.
Mutlu S, Osma E, Ilhan V, Turkoglu HI, Atici O (2016). Mistletoe (Viscum album) reduces the growth of the Scots pine by accumulating essential nutrient elements in its structure as a trap. Trees 30: 815–824.
Pilichowski S, Filip R, Koscielska A, Zaroffe G, Zyzniewska A, Iszkulo G (2018). Wpływ Viscum album ssp. austriacum (Wiesb.) Vollm. na przyrost radialny Pinus sylvestris L. Sylwan 162: 452–459. (in Polish).
R Core Team (2023). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Sanguesa-Barreda G, Linares JC, Camarero JJ (2013). Drought and mistletoe reduce growth and water-use efficiency of Scots pine. Forest Ecology and Management 296: 64–73. https://doi.org/10.1016/j.foreco.2013.01.028.
Silva FP, Fadini RF (2017). Observational and experimental evaluation of hemiparasite resistance in trees in the urban afforestation of Santarém, Pará, Brazil. Acta Amazonica 47 (4): 311–320. https://doi.org/10.1590/1809-4392201700033.
Strong GL, Bannister P (2002). Water relations of temperate mistletoes on various hosts. Functional Plant Biology 29 (1): 89–96. https://doi:10.1071/PP00159.
Szmidla H, Tkaczyk M, Plewa R, Tarwacki G, Sierota Z (2019). Impact of common mistletoe (Viscum album L.) on Scots pine forests – a call for action. Forests 10 (847): 1–15. https://doi:10.3390/f10100847.
Thomas PA, Dering M, Giertych MJ, Iszkuło G, Tomaszewski D, Briggs J (2022). Biological Flora of Britain and Ireland: Viscum album. Journal of Ecology 303: 1–39. https://doi.org/10.1111/1365-2745.14036.
Tsopelas P, Angelopoulos A, Economou A, Soulioti N (2004). Mistletoe (Viscum album) in the fir forest of Mount Parnis, Greece. Forest Ecology Management 202: 59–65. https://doi.org/10.1016/j.foreco.2004.06.032.
Türe C, Böcük H, Aşan Z (2010). Nutritional relationships between hemi-parasitic mistletoe and some of its deciduous hosts in different habitats. Biologia 65: 859–867. https://doi.org/10.2478/s11756-010-0088-5.
Turner RJ, Smith P (2016). Mistletoes increasing in eucalypt forest near Eden, New South Wales. Australian Journal of Botany 64: 171–179. https://doi.org/10.1071/BT15253.
Valle ACV, Carvalho AC, Andrade RV (2021). Viscum album – literature review. International Journal of Science and Research 10: 63–71.
Varga I, Taller J, Baltazár T, Hyvönen, J, Poczai P (2012). Leaf-spot disease on European mistletoe (Viscum album) caused by Phaeobotryosphaeria visci: a potential candidate for biological control. Biotechnology Letters 34 (6): 1059–1065. https://doi.org/10.1007/s10529-012-0867-x.
Walas Ł, Kędziora W, Ksepko M, Rabska M, Tomaszewski D, Thomas PA, Wójcik R, Iszkuło G (2022). The future of Viscum album L. in Europe will be shaped by temperature and host availability. Scientific Reports 12: 17072. https://doi.org/10.1038/s41598-022-21532-6.
Watson DM, Herring M (2012). Mistletoe as a keystone resource: an experimental test. Proceedings of the Royal Society Series B 279: 3853–3860. https://doi.org/10.1098/rspb.2012.0856.
Watson DM (2015). Disproportionate declines in ground-foraging insectivorous birds after mistletoe removal. PLoS ONE 10 (12): e0142992. https://doi.org/10.1371/journal.pone.0142992.
Wójcik R, Kędziora W (2020). Abundance of Viscum in central Poland: Results from a large-scale mistletoe inventory. Environmental Sciences Proceedings 3 (1): 7883. https://doi.org/10.3390/IECF2020-07883.
Zebec M, Idžojtić M (2006). Hosts and distribution of yellow mistletoe, Loranthus europaeus Jacq. in Croatia. Hladnikia 2 (19): 41–46.
Zar JH (2010). Biostatistical Analysis. 5th Edition, Pearson Prentice Hall, New Jersey.
Zuber D (2004). Biological flora of Central Europe: Viscum album L. Flora 199: 181–203.



