Schlüsselbegriffe: Operationscharakteristik eines Beobachters, Ellenberg-Klimaquotient, Forestry Aridity Index
Abstract
The survival of forest tree species within certain areas of their distributions is in question due to the increasing occurrence of disturbances and degradation processes in forest ecosystems due to climate change. The aim of this paper is to predict changes in the spatial distribution of the ten most important tree species in Serbia (European beech, Pedunculate oak, Austrian oak, Hungarian oak, Sessile oak, Narrow-leafed ash, Silver fir, Norway spruce, Black and Scots pine) using climate indices (Forestry Aridity Index, FAI and Ellenberg Quotient, EQ) with up-to-date climate observations (E-OBS, covering the time period 1990-2019) and projections of future climate conditions (RCP 4.5 and RCP 8.5 scenarios, split into two time periods 2041-2070, 2071-2100). The computation of the area under the receiver operating characteristic (ROC) curves has shown that both FAI and EQ have a “fair” to “excellent” ability to predict the occurrence of five out of ten species (European beech, Silver fir, Norway spruce, Black and Scots pine), with EQ having a slightly better predicting ability. EQ-based projections from mid (2041-2070) to late 21st century (2071-2100) under RCP 4.5 predict that reduction rates will not exceed 25%. Similar reduction rates are given by FAI-based projections until 2070, while rates increase to approximately 35% towards the end of this century. For RCP 8.5, FAI-based projections are significantly worse than EQ-based projections. Irrespective of the used index, projections until 2070 suggest that 55-75% of existing habitats will remain intact. Towards the end of the century, however, our analysis indicate that 75-85% (EQ) to 90-100% (FAI) of the five analyzed species habitats, will be found outside of their current climate space.
Zusammenfassung
Durch das zunehmende Auftreten von Störungen und Degradationsprozessen in Waldökosystemen aufgrund des Klimawandels ist das Überleben einiger Baumarten in bestimmten Gebieten Serbiens unklar. Das Ziel dieser Arbeit ist es, Veränderungen in der räumlichen Verteilung der zehn wichtigsten Waldbaumarten (Rotbuche, Stieleiche, Zerreiche, Ungarische Eiche, Traubeneiche, Schmalblättrige Esche, Weißtanne, Gemeine Fichte, Schwarzkiefer/Waldkiefer) in Serbien mittels Klimaindices (Forestry Aridity Index, FAI und Ellenberg-Klimaquotient, EQ) mit aktuellen Klimamessdaten (E-OBS, Zeitraum 1990-2019) und Prognosen der zukünftige Klimabedingungen (RCP 4.5 und RCP 8.5, in zwei Zeiträume geteilt, 2041-2070, 2071-2100) vorherzusagen. Die Berechnungen mittels ROC-Kurve (Operationscharakteristik eines Beobachters) zeigt, dass sowohl FAI als auch EQ "gut" bis "ausgezeichnet" geeignet sind, das Auftreten von fünf von zehn Baumarten (Rotbuche, Weißtanne, Gemeine Fichte, Schwarzkiefer/Waldkiefer) wiederzugeben, wobei EQ etwas besser geeignet war. Es zeigte sich, dass auf Basis von EQ und dem RCP 4.5 Szenario von der Mitte des 21. Jahrhunderts (2041-2070) bis zum Ende des Jahrhunderts (2071-2100) keine Reduktionsraten geeigneter Lebensräume zu erwarten sind, die 28% überschreiten. Ähnliche Reduktionsraten zeigen Prognosen des FAI bis zur Mitte des Jahrhunderts, während gegen Ende des Jahrhunderts diese auf etwa 35% steigen. Bei RCP 8.5 sind die auf FAI basierenden Projektionen im Vergleich zu EQ deutlich schlechter. Während in beiden Fällen (EQ und FAI) die Prognosen für die Mitte des Jahrhunderts vorhersagen, dass etwa 55% bis 75% der vorhandenen Lebensräume intakt bleiben wird, sind die Prognosen zum Ende des Jahrhunderts wesentlich beunruhigender. Unsere Ergebnisse zeigen, dass 75-85% (EQ) bis 90-100% (FAI) der fünf analysierten Artenlebensräume bis zum Ende des 21. Jahrhunderts außerhalb der derzeitigen klimatischen Zonen liegen werden.
1. Introduction
Climate change and its associated impacts on all aspects of life represents an enormous challenge to present and future environmental, economic, and social well-being. Forests covering 31% of the global land area (FAO and UNEP, 2020) have great social-economic and ecological importance and are particularly endangered by climate change duе to their inability to adаpt quickly to rapidly changing conditions. Trees are sensitive to climate change, as an increase of just 1 °C in mean annual air temperature is sufficient to cause significant changes in the growth and regeneration capacity of many tree species (Kirschbaum and Fischlin, 1996). The consequences of this relatively minor change have been contributed across forests in Europe in recent years to more frequent dieback, pest outbreaks, reduced productivity or loss of vitality (Spathelf et al. 2013).
Climate indices such as the Forestry Aridity Index (FAI), the Ellenberg Quotient (EQ) (Führer et al. 2011, Vlăduţ et al. 2017, Salamon-Albert et al. 2016, Nedealcov et al. 2019, Mellert et al. 2015, Trombik et al. 2013, Garamszegi and Kern, 2014, Jović et al. 2018, Móricz et al. 2018, Mátyás et al. 2018), the Standardized Precipitation Evapotranspiration Index, the Palmer Drought Severity Index, the Standardized Precipitation Index (Stojanović et al. 2015b, Boczoń et al. 2018), the Pálfai Drought Index (Ladányi and Blanka 2015, Mátyás et al. 2018), and others can be used to investigate the effects of climate on forest growth. Some of these climate indices, like FAI and EQ, as well as many bioclimatic variables, have been used in species distribution modelling (SDM) as predictors to predict future tree species occurrence on continental European scale (Lindner et al. 2014, Falk & Hempelmann 2013, Thurm et al. 2018, Buras & Menzel 2019, Noce et al. 2017, Goberville et al. 2016, Ruosch et al. 2016) and regional scales including Serbia (Pavlović et al. 2019, Stojanović et al. 2013, Stojanović et al. 2014), Montenegro (Matović, 2013), Hungary (Czúcz et al. 2011, Mátyás et al. 2018, Móricz et al. 2013), Germany (Falk & Mellert 2011, Walentowski et al. 2017), Italy (Pecchi et al. 2020, Marchi et al. 2016), Slovenia (Kutnar & Kobler, 2011), and Iran (Taleshi et al. 2019). The common denominator in each of these studies is the prediction of shifts toward higher elevations and severe reductions in climate-suitable areas of mesophilous or hygrophilous species (e.g., Scots pine, Norway spruce, Silver fir, Grand fir, European beech, Oriental beech, etc.). Some of the studies listed above, especially those on European scale, predict the sustainability of existing or expansion of climatically-suitable areas for xerophilous tree species such as Sessile oak and Black pine under climate change. Analyses conducted by Walentowski et al. (2017), Friedrichs et al. (2009), and Eilmann & Rigling (2012) suggest that there is a real threat to the survival of deciduous and coniferous species that are characterized as mesophilous and/or hygrophilous due to their poor tolerance of extreme climate events. Over the past two decades, such events have been increasingly reported for European beech (Štajner et al. 2017), Norway spruce (Matović et al. 2018, Karadžić et al. 2017), Pedunculate oak, and Austrian oak forests (Stojanović et al. 2015a, Stojanović et al. 2015b). Given that such events can be expected to occur with greater frequency in the future, SDMs can be useful for identifying indicators of degradation processes.
The aim of this study is to predict the distributions of ten tree species (Silver fir, Norway spruce, European beech, Austrian oak, Sessile oak, Pedunculate oak, Hungarian oak, Narrow-leafed ash, black and Scots pine) in Serbia using EQ and FAI indices with climate observations (E-OBS: 1990-2019) and emission scenarios (RCP 4.5 and 8.5: 2041-2070, 2071-2100).
This study represents the first attempt to model the distribution of different tree species in Serbia. The novelty in this research is the use of the most recent climate data (1990-2019), which is a significant improvement in comparison to previous studies. This study also takes advantage of the most recent climate change projection data (Representative Concentration Pathway – RCP).
2. Material and methods
2.1 Climatic data
To evaluate past and future climate characteristics, we used:
1. The forest aridity index (FAI) of Führer et al. (2011), which takes into account the average temperature of the critical months (July and August; °C) and precipitation during the main growth cycle (May to July), as well as precipitation during the critical months (July and August; mm) (1).
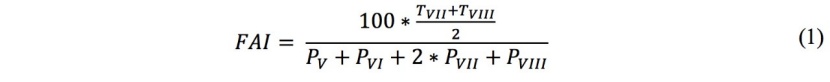
2. Ellenberg's (1988) climate quotient (EQ), which takes into account the average temperature of the warmest month (July; °C) and annual precipitation (Pannual) (2).
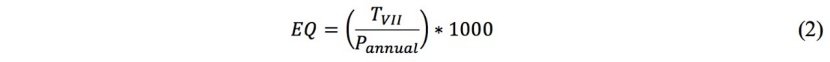
From the E-OBS v21.0 gridded dataset (Cornes et al. 2018), we extracted climatic data to calculate the FAI and EQ indices for the reference period (1990-2019) and for the past climatic sequence (1961-1990). For future projections, we used the Representative Concentration Pathway (RCP) 4.5 and 8.5 for mid- (2041-2070) and late- (2071-2100) 21st century scenarios (Marchi et al. 2020).
2.2 Spatial data analysis
Data regarding the spatial distribution of forest species were obtained from Serbia's National Forest Inventory (NFI). The inventory is based on a systematic sampling process that uses a 4x4 km network of sampling clusters (Banković et al. 2009). NFI clusters that fall outside afforested area were removed from the dataset based on satellite images. The NFI considered all inventory plots to be forested if they have an area of more than 0.5 ha and are overgrown with forest trees whose crowns cover or should reach a coverage of more than 10% of the area, where the trees must be able to reach a minimum height of 5 m at felling age. For our research, it is especially important that all trees in an inventory unit are identified and the stand affiliations estimated. Accordingly, we used rasters of stand affiliation from the NFI, due to the higher representative value of such data in terms of reading the conditionality of the occurrence of a certain species with different habitat characteristics. The selection of species for this analysis is based on the list of main tree species managed by two state enterprises for forest management – "Srbijašume" and "Vojvodinašume" (SE "Srbijašume", 2021 & SE "Vojvodinašume", 2021). Special emphasis is placed on species whose stands are predominantly of natural origin (Silver fir, Norway spruce, European beech, Austrian oak, Sessile oak, Pedunculate oak, Hungarian oak, Narrow-leafed ash, and Black and Scots pine). This eliminates possible errors that may occur due to the analysis of species whose occurrence in certain areas is the result of anthropogenic activity, such as acacia and selected black poplar and willow varieties. It is important to note that, despite their different ecological requirements, the NFI methodology registers Black and Scots pine under a single category, "pine forests". Using the R statistical environment (R Core Team, 2020) and its “base” package coupled with the “raster” package (Hijmans, 2020), the spatial distribution of forest species is converted to a binary type raster. Using packages “base” (R Core Team, 2020), “ncdf4” (Pierce, 2019), and “raster” (Hijmans, 2020), climate data are processed, converted to raster (E-OBS), resampled, cropped to match resolution and extent of the binary type raster, and exported to an ArcMap 10.5 (ESRI, 2016) supported format. Maps of elevation and total forest cover were created within ArcMap 10.5 using the European Digital Elevation Model (EU-DEM) version 1.1. and Corine Land Cover (CLC) 2018 data from the European Environment Agency (EEA).
To examine whether FAI and EQ indices can predict the occurrence of the analyzed species, we computed the area under the receiver operating characteristic (AUC), a widely used statistic for assessing the discriminatory capacity of species distribution models (Jiménez-Valverde, 2011), using the R package “pROC” (Robin et al. 2020). Accuracy classification of AUC has been carried out using a scale where: 0.50–0.60 = fail; 0.60–0.70 = poor; 0.70–0.80 = fair; 0.80–0.90 = good; 0.90–1 = excellent (Swets 1988, as cited in Rasztovits 2011). Bearing in mind that algorithms with low AUC values produce very inconsistent spatial predictions (Aguirre-Gutiérrez et al. 2013), we have omitted species with “poor” accuracy from the prediction analysis. To determine best-fitting FAI and EQ values that describe the occurrence of the analyzed species, we used a threshold that maximizes the sum of sensitivity and specificity (Cantor et al. 1999, as cited in Liu et al. 2005, Czúcz et al. 2011, as cited in Stojanović et al. 2013).
With the help of the extract function from the “raster” package (Hijmans, 2020), we determined the minimum, maximum, mean, and standard deviation of FAI and EQ indices for the distribution of each current forest type (excluding the autonomous province of Kosovo and Metohija) in the reference and past climate sequences. We also calculated the differences between the mean values of the FAI and EQ indices of both climate sequences. Based on the previously calculated thresholds for each species, the FAI and EQ rasters for the reference and projected periods were reclassified to consist of two classes using the “base” (R Core Team, 2020) and “raster” (Hijmans, 2020) packages. The first class contains all raster pixels whose values are equal to or less than the specified threshold; all other pixels make up the second class. The relative share of the total number of raster pixels in the first class represents the relative share of climatically-suitable habitats in the observed and projected periods. These calculations were performed separately for the area of current-day forests (Table 3) and the total area of Serbia (Table 4). The relative difference (Δ) between the values for the observed and projected periods represents a gain or loss of climatically-suitable habitats depending on whether the difference is positive or negative. It is important to note that the relative share of climatically-suitable habitats within areas of current-day forests in the reference period also represents an indicator of threshold spatial precision (Table 3, column E-OBS). The final map visualization was completed using ArcGIS 10.5 software (ESRI, 2016).
3. Results and Discussion
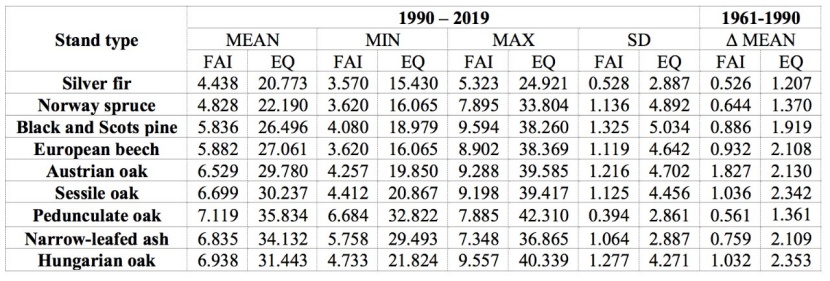
Table 1: Minimum, maximum, mean, and standard deviations of FAI and EQ values in current forest distribution. / Tabelle 1: Minimale, maximale, mittlere und Standardabweichung der FAI- und EQ-Werte in der aktuellen Waldverteilung.
According to FAI and EQ values (Table 1), Hungarian oak forests and floodplain forests of Narrow-leaved ash and Pedunculate oak occur in the most ''arid'' zone. Other oaks also occupy more arid habitats, whereas Silver fir and Norway spruce grow in the most humid climates. European beech and Black and Scots pine are found in areas characterized by a moderately humid climate. In accordance with their growth and development capabilities, Austrian oak, Sessile oak, Hungarian oak, European beech, and Black and Scots pines are found in a wide range of climatic conditions (Table 1). As expected, species referred to as “azonal” (Narrow-leafed ash and Pedunculate oak) and “climax” (Silver fir and Norway spruce) have the narrowest ecological niches (Table 1). Many such species constitute a significant areas of forests, especially European beech in hilly and mountainous areas (Pavlović et al. 2019) and pedunculate oak and narrow-leaved ash in lowlands (Ivanišević and Knežević, 2008, as cited in Bobinac et al. 2010).
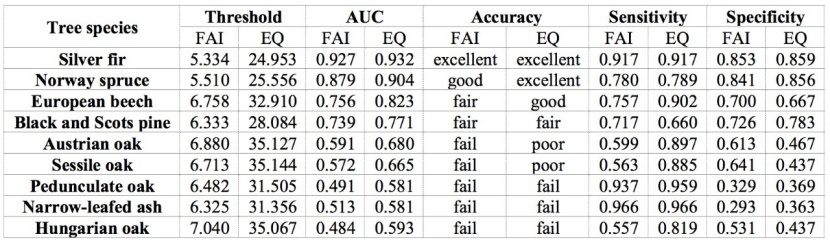
Table 2: Statistical parameters for FAI and EQ index based on E-OBS observed climate dataset (1990 – 2019). / Tabelle 2: Statistische Parameter für FAI- und EQ-Index basierend auf dem von E-OBS beobachteten Klimaangaben (1990 – 2019).
Compared to the previous climate sequence (1961-1990), the trend of increasing FAI values in the more recent sequence is least pronounced in areas where Silver fir and Norway spruce are present (Table 1). This is expected due to their dominance at higher elevations (Figure 1), where warmer temperatures are more likely to benefit growth. Interestingly, lowland forests of Pedunculate oak and Narrow-leafed ash also exhibit minor FAI trends. In the context of this study, increase of FAI values in areas where the most widespread tree species is present - European beech, can be described as moderate. Almost the same trend is observed in the areas populated by oaks (except for Austrian oak) and pines (Table 1). These extreme results for the widespread Austrian oak are not surprising (Table 1), as it inhabits predominantly xerophilous habitats at lower elevations (Figure 1). These habitats are especially vulnerable to climate change. In the case of the EQ index, the differences between the two climate series are fairly uniform among all species, except for areas inhabited by Silver fir, Norway spruce and Pedunculate oak, where the differences are the least pronounced (Table 1). Nevertheless, their narrow ecological niche makes them highly vulnerable to climate change. Interestingly, our results (Table 1) show that some European beech stands (Figure 1) are found close to the marking value (EQ=40) for its disappearance (Budeanu et al. 2016, as cited in Vlăduţ et al. 2017). Given to the observed climatic trend, the number of stands that will be close to this value will only increase in the coming years.
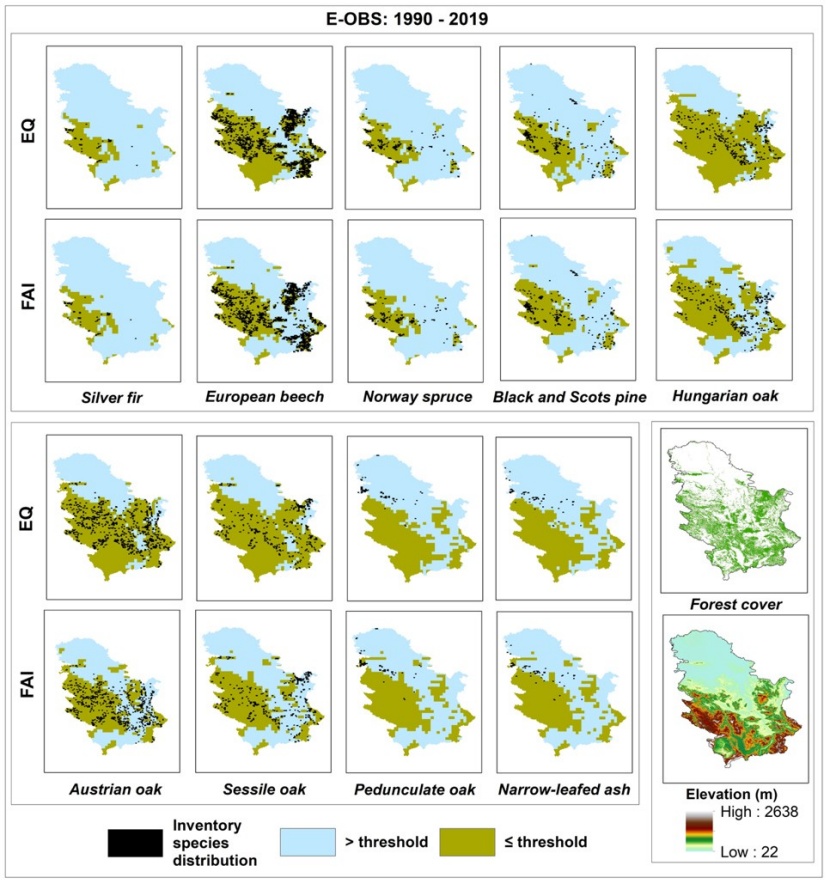
Figure 1: Map of the current forest trees distribution with indicated FAI and EQ index thresholds for observed climate (E-OBS, 1990-2019) with the display of elevations and overall forest cover in Serbia. / Abbildung 1: Karte der aktuellen Waldbaumverteilung mit den angegebenen FAI- und EQ-Indexschwellenwerten für das untersuchte Klima (E-OBS, 1990-2019) mit Anzeige der Höhen und der gesamten Waldbedeckung in Serbien.
3.1 Determination of the predictive accuracy of FAI and EQ
According to our analysis, the FAI index has a “fair” to “excellent” ability to discriminate the occurrence of four out of ten analyzed species, with AUC ranging from 0.739 to 0.927 (Table 2). In contrast, the EQ index has a “poor” to “excellent” ability to discriminate the occurrence of six out of nine analyzed species, with AUC ranging from 0.665 to 0.932 (Table 1). The lowest AUC values in both indices are mainly found in azonal species (e.g., Pedunculate oak and Narrowed-leafed ash) and species with wide ecological niches (e.g., oaks). For this reason, they are omitted from further analysis. As expected, the highest AUC values are found in climax species (e.g., Silver fir and Norway spruce). FAI thresholds show different spatial precision (Table 4) that is usually highly correlated with AUC accuracy (Table 2). This is not the case with Norway spruce, however. Although the accuracy of its AUC value is classified as “good”, the Norway spruce threshold describes only two-thirds (78.08%) of its spatial distribution (Table 3 and Figure 1.). Similar results are obtained for Black and Scots pine, but to a lesser extent. Norway spruce has a “fair” AUC classification and its EQ threshold describes 65.89% of its current-day distribution. Such results should not be surprising. According to Tomić et al. (2011), forest plantations of Norway spruce, and Black and Scots pine were mass-produced between the 1950s and the 1970s in areas with inadequate habitat characteristics. The low precision of the pine AUCs can also be explained by the different ecological needs of Black and Scots pine, which are lumped together in the NFI classification. However, the spatial precision of the EQ thresholds are quite similar to those of the FAIs, with one exception (Table 3 and Figure 1). The EQ threshold explains almost all (90.23%) of the spatial distribution of European beech, in contrast to its FAI analogue (75.99%).
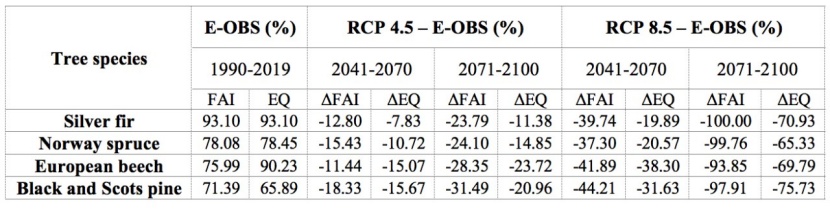
Table 3: Percent of climatically-suitable areas and relative difference between observed and projected climate conditions within current NFI trees distribution based on FAI and EQ index and their threshold. / Tabelle 3: Prozentsatz klimatisch geeigneter Gebiete und relativer Unterschied zwischen beobachteten und projizierten Klimabedingungen innerhalb der aktuellen Baumverteilung basierend auf FAI- und EQ-Index und deren Schwellenwert.
3.2 FAI- and EQ-based projections
The increased FAI and EQ values shown in Figures 2 and 3 indicate that parts of Serbia, especially the southern and southeastern parts, are likely to be severely impacted by climate change. However, local and international studies indicate that the reactions of forests to climate change are likely to differ depending on the overall ecological needs and provenance characteristics of their species (Stojanović et al. 2015a, Mikac et al. 2018, Horváth and Mátyás 2016, Matović et al. 2018, Rybníček et al. 2012, Čermák et al. 2017, Castagneri et al. 2015, Tikvić et al. 2008, Jović et al. 2018, Gavrilov et al. 2019, Horák et al. 2014, Stjepanović et al. 2017, Carrer et al. 2012, Spathelf et al. 2013, Führer et al. 2016, Isaac-Renton et al. 2018, George et al. 2015, Taeger et al. 2013, Arend et al. 2011, Robson et al. 2012, Rose et al. 2009, Matías et al. 2016). This should be kept in mind because increasing aridity levels do not explicitly mean the disappearance of species. Further investigation into the impacts of climate change could tell us much more about current and future environmental processes.
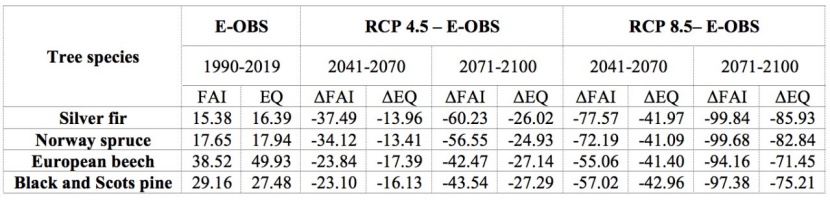
Table 4: Percent of climatically-suitable areas and relative difference between observed and projected climate conditions within whole territory of Serbia based on FAI and EQ index and their threshold. / Tabelle 4: Prozentsatz klimatisch geeigneter Gebiete und relativer Unterschied zwischen beobachteten und projizierten Klimabedingungen innerhalb des gesamten Territoriums Serbiens basierend auf FAI- und EQ-Index und deren Schwellenwert.
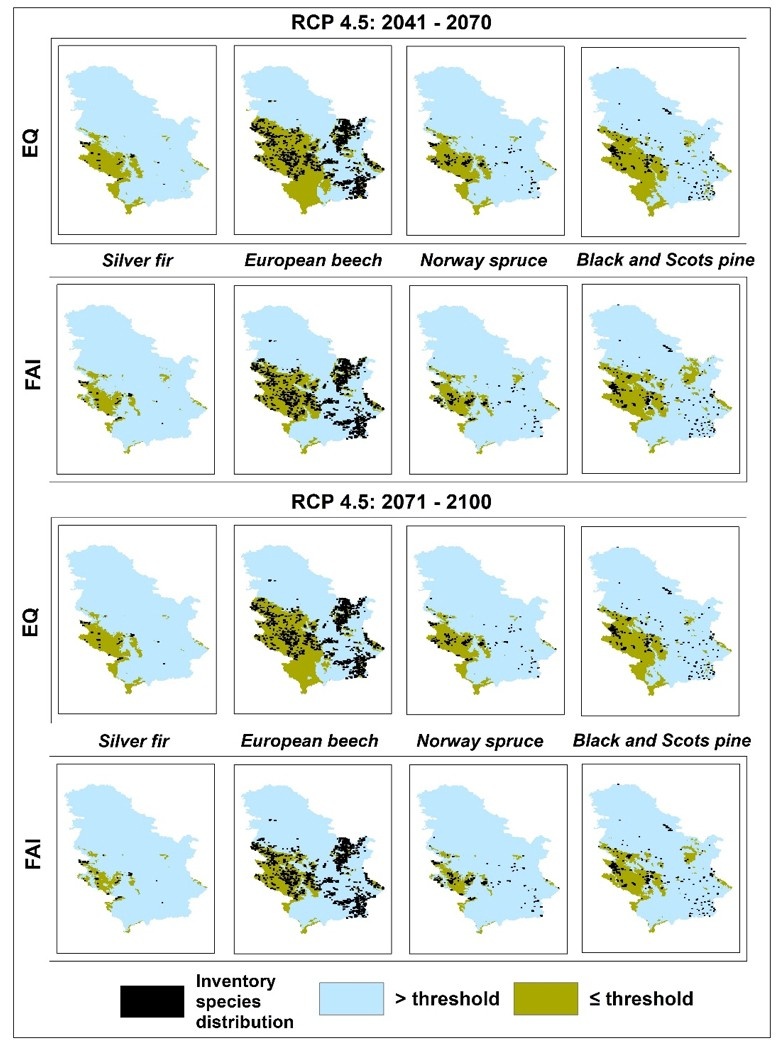
Figure 2: Map of the current forest tree species distribution over FAI and EQ maps for projected climate (RCP 4.5 for periods 2041-2070 and 2071-2100). / Abbildung 2: Karte der aktuellen Verteilung der Waldbaumarten über FAI- und EQ-Karten für das projizierte Klima (RCP 4.5 für die Zeiträume 2041-2070 und 2071-2100).
3.2.1 Silver fir and Norway spruce
According to FAI- and EQ-based RCP 4.5 projections, climatically-suitable areas throughout Serbia, where the presence of Silver fir and Norway spruce is not recorded, will partially withdraw from their eastern and southern "borders" towards higher elevations in the western part of Serbia by mid-century (2041-2070). Late-century (2071-2100) projections predict an intensification of these changes (spatially and value-wise). Up to 2.5 times higher reduction rates under FAI-based projections than those under EQ-based projections (Table 4) results in the narrowing of climate-friendly areas around existing spruce and fir habitats (Figure 1 and 2). Since FAI provides more extreme predictions, it may be worth stressing that it has stronger focus on climate data during growing season. Thus, Silver fir and Norway spruce may not be particularly endangered, as both EQ- and FAI-based projections give similar reduction rates that are relatively small and constant from mid to late century (around 20%) (Table 3). According to these projections, existing Norway spruce sites in the southeast and few sites of Silver fir in the east will be found under the impact of climate change (Figure 1 and 2).
FAI- and EQ-based mid-century projections under RCP 8.5 predict spatial changes similar to those under the RCP 4.5 scenario, but with almost twice the reduction rate of suitable habitats (Table 4). In this case, the withdrawal of suitable habitats is most pronounced from the northeast to the southwest. (Figures 1, 2, and 3). Such changes will increase the reduction rates of suitable habitats for Silver fir and Norway spruce by 20.96% (EQ) to 31.49% (FAI) (Table 3). Late-century FAI- and EQ-based projections under RCP 8.5 are even less promising, as they predict the severe to complete reduction of suitable habitats. The same is seen for the entire territory of Serbia (Table 4). Only the EQ-based projections leave some room for the survival of Silver fir and Norway spruce at higher elevations in west Serbia (Table 3, Figures 1 and 3).
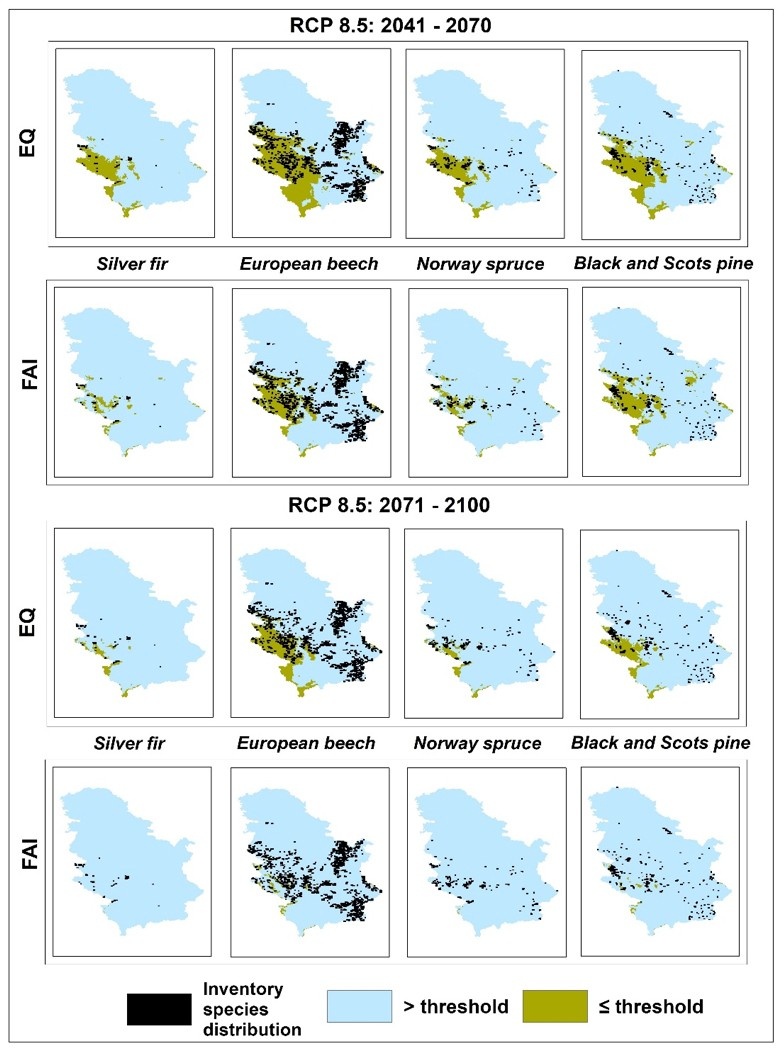
Figure 3: Map of the current forest tree species distribution over FAI and EQ maps for projected climate (RCP 8.5 for periods 2041-2070 and 2071-2100). / Abbildung 3: Karte der aktuellen Verteilung der Waldbaumarten über FAI- und EQ-Karten für das projizierte Klima (RCP 8.5 für die Zeiträume 2041-2070 und 2071-2100).
3.2.2 European beech
In the case of European beech, all of the FAI index projections are quite uniform (Tables 3 and 4). From the mid to late century, they all predict an increasing reduction of climatically-suitable habitats (Table 4) toward the west, especially at lower elevations where the presence of European beech is not currently recorded (Figures 1, 2, and 3). In contrast, all major changes under EQ-based projections are those affecting the spatial distribution of European beech (Figures 1, 2, and 3). These predictions are to be expected as the threshold obtained from the ROC analysis almost perfectly outlines the distribution of European beech in Serbia. With regard to the distribution of European beech, FAI- and EQ-based projections under RCP 4.5 give almost identical results, predicting a slight reduction of suitable habitats at mid-century (not exceeding 15%), with only a slight further decrease by the end of the century (Table 3), especially in the south and southeast (Figures 1 and 2).
Similarly, there is little difference in the FAI- and EQ-based mid-century projections of the distribution of European beech under RCP 8.5. Both projections predict a severe reduction of suitable habitats in the south and southeast and, to a lesser extent, in the east and west (Figures 1 and 3). Compared to their RCP 4.5 analogues, however, the overall reduction rates are up to three or four times greater (Table 3). The EQ-based late-century projection gives more promising results as compared to the almost complete reduction predicted by its FAI analogue. The EQ-based projections predicts that 30.21% of European beech (Table 3) distributed at higher elevations in west Serbia may remain intact (Figures 1 and 3).
3.2.3 Black and Scots pine
According to the RCP 4.5 mid-century FAI- and EQ-based projections, climatically-unsuitable areas for Black and Scots pine in Serbia will expand by about 20% (Table 4) in the central part of the country (in the form of several "peninsulas"). Such changes will be largely present in directions from the east to the west and from the south to the north, exclusively at lower elevations where the presence of Black and Scots pine has not been recorded (Figures 1 and 2). However, an expansion of climatically-suitable areas under the FAI-based projection is predicted in the east, where the presence of Black and Scots pine has been recorded in smaller areas (Figures 1 and 2). In the south and southeast, where pine is more common, both FAI- and EQ-based projections predict reductions of almost 100%. This contributes significantly to the overall reduction of suitable habitats in Black and Scots pine inhabited areas (which is around 15%) (Table 3, Figures 1 and 2). FAI- and EQ-based RCP 4.5 projections for the late 21st century predict almost twice the reduction rate as compared to mid-century projections (Table 4). The expansion of climatically-unsuitable areas is predicted further on the directions observed in mid-century projections, whereby they will partially enter into the areas where Black and Scots pine are present. (Figures 1 and 2). With regard to the distribution of Black and Scots pine, common in both (FAI- and EQ-based) projections is a relatively small increase of reduction rates, compared to those from mid-century projections (Table 3). This is especially pronounced in the case of EQ-based projection (Table 3). Interestingly, the increase in climatically-suitable areas in eastern Serbia predicted by the FAI-based mid-century projection under RCP 4.5 now decreases to the state in the observed climatic period (E-OBS: 1991-2019) (Figures 1 and 2).
Mid-century projections under FAI- and EQ-based RCP 8.5 predict a complete reduction of suitable habitats in the east and southeast, which makes up about half of the total area of suitable habitats in Serbia (Table 4), leaving only the western distributions of Black and Scots pine intact (Table 3, Figures 1 and 3). Аs usual, the FAI-based projection (under RCP 8.5) predicts a slightly higher reduction rate (Table 3), resulting in the expansion of unsuitable habitats in southwestern Serbia (Figures 1 and 3). In contrast to the FAI-based late-century projection under RCP 8.5, which predicts an almost complete reduction of suitable habitats, the EQ-based projection intensifies changes (spatially and value-wise) predicted by the mid-century projection. However, it still leaves some room for the survival of 24.27% of Black and Scots pine sites located in the higher elevations in the west (Tables 3 and 4, Figures 1 and 3).
3.3 Comparison and perspectives
Climate change in Serbia is already a reality; its consequences are expected to become more intense in the coming decades. Meteorological measurements provided by the Republic Hydrometeorological Service of the Republic of Serbia for the last several decades (1961-1990 and 1981-2010) point to an increasingly changed climatic situation. Several meteorological stations in western Serbiа (Kopaonik, Zlatibor, Sjenica) have recorded a slight increase in temperature (≈0.5°C), which has been accompanied by an increase in precipitation of about 5% on an annual basis. These relatively moderate climate changes can positively affect forest productivity (Gustafson et al. 2017). In contrast, the same pattern of temperature increase is accompanied by a decrease in precipitation of approximately 5% in the eastern (Crni vrh, Negotin, Zajecar) and southeastern (Dimitrovgrad, Leskovac, Vranje) parts of Serbia. In this case, trees struggle to maintain productivity because the increased level of transpiration caused by the increase in temperature needs to be accompanied by sufficient precipitation (Kirschbaum 2000). In both cases, the timing of precipitation shifts noticeably from the vegetation period to the winter period, with significant consequences for the forests. For example, Matović et al. (2018) found that extremely high temperatures during the vegetation period in combination with extremely low rainfall is the primary cause of devitalization and dieback of spruce trees. Spatial distribution of these (positive and negative) trends of climate change, observed by RHMS of Serbia, fully complies with the shift pattern of climate suitable habitats predicted by RCP 4.5 and especially RCP 8.5. The strong retreat of those habitats toward higher elevations in the west (Figures 1 and 2), predicted by the mentioned scenarios, is in line with the results of other SDM studies listed in the introduction. With regard to reduction rates, we found no reduction rate (over 35%) under any of the RCP 4.5 and RCP 8.5 EQ-based mid-century projections that would significantly jeopardize the survival of tree species habitats in Serbia as we know them today (Table 3). Surprisingly, FAI- and EQ-based projections under RCP 4.5 do not deviate significantly from one another. FAI-based projections under RCP 8.5 give much greater reduction rates than their EQ analogues, but only for species with narrow ecological niches (Silver fir and Norway spruce). These differences also exist in late-century projections, but include all species (Table 3).
3.3.1 European beech
Projections under RCP 8.5, especially those based on FAI (Table 3), are in line with the results of Kramer et al. (2010), which also predict that beech habitats in Greece and ex-Yugoslavia will become climatically-unsuitable by 2050. According to Saltré et al. (2014), this change is likely to lead to an increase in drought mortality of trees in 2081-2100 in these regions. Several other studies of European beech in Serbia (Stojanović et al. 2013), Hungary (Móricz et al. 2013, Czúcz et al. 2011), and Slovenia (Kutnar & Kobler 2011) have come to similar conclusions; all are in agreement that almost all beech stands will be found outside climatically-suitable zones toward the end of the century. To maintain beech populations at existing sites, non-resistant populations should be gradually replaced by resistant ones with appropriate management measures. Particular attention should be given to populations of European beech at lower elevations due to their high vulnerability to drought events (Stjepanović et al. 2017). A study by Horváth & Mátyás (2016) in which the translocation of beech populations from different regions of Europe to the test site were used to mimic responses to climate change highlights the importance of using planting stock that is suited to the future climate conditions at the planting site.
3.3.2 Silver fir and Norway spruce
Recent research on Silver fir (Klopčić et al. 2017, Mina et al. 2015, Falk & Mellert 2011, Ruosch et al. 2016) and Norway spruce (Buras and Menzel 2019, Mina et al. 2015) predict significant reductions in their relative abundance in existing forests or even their complete disappearance. Our study and others point to significant future declines in the abundance of Silver fir (Ruosch et al. 2016) and Norway spruce (Buras & Menzel 2019) under RCP 8.5, particularly in the southern parts of Europe. Given that today’s southern European Silver fir forests are confined to moist and cool habitats at intermediate to high elevations (Ruosch et al. 2016) and that forests of Norway spruce have already been raised successfully in warm localities beyond its natural distribution range (Falk & Hempelmann 2013), there is a chance that these tree species will continue to survive in their existing habitats. However, it should be noted that fir is quite sensitive to environmental (climate) variability (Carrer et al. 2012), which suggests that any minor changes in climate are likely to significantly affect its growth. If climate-induced degradation or extinction of existing Silver fir and/or Norway spruce habitats occurs, it may be possible to compensate for the loss by introducing these species to climatically-appropriate areas at higher elevations, especially in beech dominated western Serbia. Such stands may develop naturally as a result of climate change; the simulations of Mina et al. (2015) for high elevations in the Dinaric Mountains of Slovenia indicate a transition from typical upper montane beech-dominated forest stands to mixed beech–fir–spruce stands. These processes should be supported by appropriate management measures because the mixing of Norway spruce and/or Silver fir with European beech improves the overall resistance and stability of the forest, increasing its resilience to unfavourable climate conditions (Falk & Hempelmann 2013, Paul et al. 2019, Neuner et al. 2014, Schwarz & Bauhus 2019). To adapt to the expected reduction in productivity, Lindner et al. (2014) point to the successful experience of introducing Douglas fir as a replacement for spruce in certain parts of Europe. From an economic point of view, this type of adaptation is more than satisfactory. However, the potential environmental consequences of such a management decision should be considered with care to ensure the sustainability and multifunctionality of the resulting forest.
3.3.3 Black and Scots pine
Although, compared to other species, projections under RCP 4.5 and RCP 8.5, especially those based on FAI, predict the highest reduction rates of Black and Scots pine suitable habitats (Table 3), one possible option for maintaining their populations is to establish forest plantations in areas that remain climatically suitable. Conceptual solutions are already available based on previous experiences with raising such forests. Tomić et al. (2011), for example, recommend that Scots pine plantations be raised in the zone of mixed forests of beech, fir, and/or spruce above 1200 m, although that may not be true for future climate conditions. This represents the upper limit at which Black pine used to grow in Serbia, whereby its introduction into the area of Sessile oak forests (with Hornbeam) to mountain beech forests is recommended (Tomić et al. 2011). In essence, habitats that pine trees are unable to make full use of should be avoided (Stojanović & Krstić 2009). It is important to note that the above-mentioned guidelines should be indicative, as climate change may give a significant contribution to habitat change in a vertical sense, due to the more frequent occurrence of summer dry periods at lower elevations. Such conditions can negatively affect the growth of Black pine, which are known to be sensitive to summer drought events, particularly at lower elevations (Hungary: Móricz et al. 2018; Turkey: Doğan & Köse 2019; Serbia: Ćirković-Mitrović et al. 2013, Koprivica et al. 2009; Spain: Herrero et al. 2013). This is also true of Scots pine according to studies in Spain (Bogino et al. 2009, Herrero et al. 2013), Poland and Hungary (Misi et al. 2019), Bulgaria (Panayotov et al. 2012), and Romania (Sidor et al. 2019).
4. Conclusion
In summary, this study and others make clear that forests will change in response to climate change, but the exact nature and extent of these changes are not yet well understood. Continued monitoring and experimentation are needed to ensure that appropriate management measures are taken to mitigate or eliminate these changes and to safeguard the health of forest ecosystems.
Acknowledgment
We acknowledge Ministry of Education, Science and Technological Advancement of Republic of Serbia (No. 451-03-68/2020-14/ 200197 and 451-03-9/2021-14/ 200197)
Reference
Aguirre-Gutiérrez, J., Carvalheiro, L. G., Polce, C., van Loon, E. E., Raes, N., Reemer, M., & Biesmeijer, J. C. 2013. Fit-for-Purpose: Species Distribution Model Performance Depends on Evaluation Criteria – Dutch Hoverflies as a Case Study. PLoS ONE, 8(5).
Arend, M., Kuster, T., Gunthardt-Goerg, M. S., & Dobbertin, M. 2011. Provenance-specific growth responses to drought and air warming in three European oak species (Quercus robur, Q. petraea and Q. pubescens). Tree Physiology, 31(3), 287–297.
Banković, S., Medarević, M., Pantić, D., Petrović, N. (Eds.) 2009. National Forest Inventory of the Republic of Serbia. [in Serbian] Ministry of Agriculture, Forestry and Water Management of the Republic of Serbia, Forest Directorate, Belgrade.
Bobinac, M. Andrašev, S., Šijačić-Nikolić, M. 2010. Elements of growth and structure of narrow-leaved ash (Fraxinus angustifolia Vahl) annual seedlings in the nursery on fluvisol, Periodicum Biologorum, 112(3): 341–351.
Boczoń, A., Kowalska, A., Ksepko, M., & Sokołowski, K. 2018. Climate Warming and Drought in the Bialowieza Forest from 1950–2015 and Their Impact on the Dieback of Norway spruce Stands. Water, 10(11), 1502: 1–18.
Bogino S., Fernández Nieto M.J., Bravo F. 2009. Climate effect on radial growth of Pinus sylvestris at its southern and western distribution limits. Silva Fennica, 43(4).
Budeanu M., Petritan A. M., Popescu F., Vasile D., Tudose N. C. 2016. The Resistance of European Beech (Fagus sylvatica) from the Eastern Natural Limit of Species to Climate Change. Notulae Botanicae Horti Agrobotanici. Universitaria Press. Cluj-Napoca. 44(2): 625-633.
Buras, A., & Menzel, A. 2019. Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios. Frontiers in Plant Science, 9, 1–13.
Cantor, S. B. et al. 1999. A comparison of C/B ratios from studies using receiver operating characteristic curve analysis. - J. Clin. Epidemiol. 52: 885-892.
Carrer, M., Motta, R., & Nola, P. 2012. Significant Mean and Extreme Climate Sensitivity of Norway spruce and Silver Fir at Mid-Elevation Mesic Sites in the Alps. PLoS ONE, 7(11), e50755: 1–9.
Castagneri, D., Petit, G., & Carrer, M. 2015. Divergent climate response on hydraulic-related xylem anatomical traits of Picea abies along a 900-m altitudinal gradient. Tree Physiology, 35(12): 1378–1387.
Čermák, P., Rybníček, M., Žid, T., Andreassen, K., Børja, I., Kolář, T. 2017. Impact of climate change on growth dynamics of Norway spruce in south-eastern Norway. Silva Fennica, 51(2): 1–16.
Ćirković-Mitrović, T., Popović, V., Brašanac-Bosanac, Lj., Rakonjac Lj., And Lučić, A. 2013. The Impact of Climate Elements on the Diameter Increment of Austrian Pine (Pinus nigra Arn.) In Serbia, Arch. Biol. Sci., Belgrade, 65(1), 161-170, 2013.
Cornes, R., G. van der Schrier, E.J.M. van den Besselaar, and P.D. Jones. 2018. An Ensemble Version of the E-OBS Temperature and Precipitation Datasets, J. Geophys. Res. Atmos., https://www.ecad.eu/ (access date, 17 June, 2020).
Czúcz, B., Gálhidy, L., & Mátyás, C. 2011. Present and forecasted xeric climatic limits of beech and sessile oak distribution at low altitudes in Central Europe. Annals of Forest Science, 68(1): 99–108.
Doğan, M., & Köse, N. 2019. Influence of Climate on Radial Growth of Black Pine on the Mountain Regions of Southwestern Turkey. Plants, 8(8), 276.
Eilmann, B., & Rigling, A. 2012. Tree-growth analyses to estimate tree species’ drought tolerance. Tree Physiology, 32(2), 178–187.
Ellenberg, H. 1988. Vegetation Ecology of Central Europe, fourth ed. Cambridge University Press, Cambridge.
ESRI. 2016. ArcGIS Desktop: Release 10.5 Redlands, CA: Environmental Systems Research Institute.
European Union (EU), Copernicus Land Monitoring Service 2018. European Environment Agency (EEA): EU-DEM v1.1. https://land.copernicus.eu/imagery-in-situ/eu-dem (access date, 11 October, 2019).
European Union (EU), Copernicus Land Monitoring Service 2018. European Environment Agency (EEA): CLC 2018. https://land.copernicus.eu/pan-european/corine-land-cover/clc2018 (access date, 02 November, 2019).
Falk, W. & Hempelmann, N. 2013. Species Favourability Shift in Europe due to Climate Change: A Case Study for Fagus sylvatica L. and Picea abies (L.) Karst. Based on an Ensemble of Climate Models, Hindawi Publishing Corporation, Journal of Climatology, Volume 2013, 1–18.
Falk, W., & Mellert, K. H. 2011. Species distribution models as a tool for forest management planning under climate change: risk evaluation of Abies alba in Bavaria. Journal of Vegetation Science, 22(4), 621–634.
FAO and UNEP. 2020. The State of the World’s Forests 2020. In brief. Forests, biodiversity and people. Rome.
Friedrichs, D. A., Trouet, V., Büntgen, U., Frank, D. C., Esper, J., Neuwirth, B., & Löffler, J. 2009. Species-specific climate sensitivity of tree growth in Central-West Germany. Trees, 23(4), 729–739.
Führer, E., Edelényi, M., Horváth, L., Jagodics, A., Jereb, L., Kern, Z., Moring, A., Szabados, I., Pödör, Z. 2016. Effect of weather conditions on annual and intra-annual basal area increments of a beech stand in the Sopron Mountains in Hungary. Idojaras – Quarterly Journal of the Hungarian Meteorological Service, 120(2): 127-161.
Führer, E., Horváth, L., Jagodics, A., Machon, A., Szabados, I. 2011. Application of new aridity index in Hungarian forestry practice. Időjárás 115(3): 205-216.
Garamszegi, B., & Kern, Z. 2014. Climate influence on radial growth of Fagus sylvatica growing near the edge of its distribution in Bükk Mts., Hungary. Dendrobiology, 72: 93–102.
Gavrilov, M. B., An, W., Xu, C., Radaković, M. G., Hao, Q., Yang, F., Guo, Z., Perić, Z., Gavrilov, G., Marković, S. B. 2019. Independent Aridity and Drought Pieces of Evidence Based on Meteorological Data and Tree Ring Data in Southeast Banat, Vojvodina, Serbia. Atmosphere, 10(10), 586: 1–19.
George, J.-P., Schueler, S., Karanitsch-Ackerl, S., Mayer, K., Klumpp, R. T., & Grabner, M. 2015. Inter- and intra-specific variation in drought sensitivity in Abies spec. and its relation to wood density and growth traits. Agricultural and Forest Meteorology, 214-215, 430–443.
Goberville, E., Hautekèete, N. C., Kirby, R. R., Piquot, Y., Luczak, C., & Beaugrand, G. 2016. Climate change and the ash dieback crisis. Scientific Reports, 6(1):1–9.
Gustafson J. E., Miranda R. B., De Bruijn M. G. A., Sturtevant R. B., Kubiske E. M. 2017. Do rising temperatures always increase forest productivity? Interacting effects of temperature, precipitation, cloudiness and soil texture on tree species growth and competition, Environmental Modelling & Software, 97: 171–183.
Herrero, A., Rigling, A., & Zamora, R. 2013. Varying climate sensitivity at the dry distribution edge of Pinussylvestris and P. nigra. Forest Ecology and Management, 308, 50–61.
Hijmans, J. R., 2020. raster: Geographic Data Analysis and Modeling. R package version 3.0-12.
Horák, R., Borišev, M., Pilipović, A., Orlović, S., Pajević, S., Nikolić, N. 2014. Drought impact on forest trees in four nature protected areas in Serbia, Šumarski list, 5–6 (2014): 301–308.
Horváth, A., Mátyás C. 2016. The Decline of Vitality Caused by Increasing Drought in a Beech Provenance Trial Predicted by Juvenile Growth. South-east Eur for, 7(1): 21–28.
Isaac-Renton, M., Montwé, D., Hamann, A., Spiecker, H., Cherubini, P., & Treydte, K. 2018. Northern forest tree populations are physiologically maladapted to drought. Nature Communications, 9(1).
Ivanišević P, Knežević M., 2008. Types of forests and forest land in the area of Ravni Srem, In: Tomović, Z. (ed) 250 years of Ravni Srem Forestry. State Enterprise “Vojvodinašume” – Forest holding Sremska Mitrovica, p. 87–118 [in Serbian].
Jiménez-Valverde, A. 2011. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Global Ecology and Biogeography, 21(4): 498–507.
Jović, G., Dukić V., Stajić, B., Kazimirović, M., Petrović D. 2018. A dendroclimatological analysis of fir (Abies alba Mill.) growth in the Borja mountain area of Bosnia and Herzegovina, Bulletin of the Faculty of Forestry, 2018 (118): 27–45.
Karadžić, D., Milanović, S., Golubović Ćurguz, V. 2017. Causes of spruce (Picea abies Karst.) dieback in the area of the nature park "Golija" [in Serbian], University of Belgrade, Faculty of Forestry: 15–17, Belgrade.
Kirschbaum, M. & Fischlin, A., 1996. Climate change impacts on forests. In: Watson, R., Zinyowera, M.C. & Moss, R.H. (eds.), Climate change 1995 – Impacts, adaptations and mitigation of climate change: scientific-technical analysis. Contribution of Working Group II to the Second Assessment Report of the Intergovernmental Panel of Climate Change (IPCC). Cambridge University Press, Cambridge a.o., pp. 95–129.
Kirschbaum, M. U. F. (2000). Forest growth and species distribution in a changing climate. Tree Physiology, 20(5-6), 309–322.
Klopčić, M., Mina, M., Bugmann, H., & Bončina, A. 2017. The prospects of Silver fir (Abies alba Mill.) and Norway spruce (Picea abies (L.) Karst) in mixed mountain forests under various management strategies, climate change and high browsing pressure. European Journal of Forest Research, 136(5-6), 1071–1090.
Koprivica, M., Matović, B., Stajić, S., Čokeša, V. 2009. Diameter increment trend of the Austrian pine plantations in Raška, Sustainable Forestry, Collection 59–60, 2009.
Kramer, K., Degen, B., Buschbom, J., Hickler, T., Thuiller, W., Sykes, M.T. and de Winter, W. 2010. Modelling exploration of the future of European beech (Fagus sylvatica L.) under climate change – range, abundance, genetic diversity and adaptive response. Forest Ecology and Management 259, 2213-2222.
Kutnar, L. & Kobler, A. 2011. Prediction of forest vegetation shift due to different climate-change scenarios in Slovenia, Šumarski list br. 3–4, CXXXV (2011), 113–126.
Ladányi Z. & Blanka V. 2015. Tree-ring width and its interrelation with environmental parameters: case study in central Hungary, Journal of Environmental Geography 8 (3–4), 53–59.
Lindner, M., Fitzgerald, J. B., Zimmermann, N. E., Reyer, C., Delzon, S., van der Maaten, E., Schelhaas M., Lasch P., Eggers J., j , van der Maaten-Theunissen M., Suckow F., Psomas A., Poulter B., Hanewinkel, M. 2014. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? Journal of Environmental Management, 146, 69–83.
Liu, C., Berry, P. M., Dawson, T. P., & Pearson, R. G. 2005. Selecting thresholds of occurrence in the prediction of species distributions. Ecography, 28(3): 385–393.
Marchi, M., Nocentini, S. and Ducci, F. 2016. Future scenarios and conservation strategies for a rear-edge marginal population of Pinus nigra Arnold in Italian central Apennines, Forest Systems 25(3), e072, 12 pages (2016).
Marchi, M., Castellanos-Acuna, D., Hamann, A., Wang, T., Ray, D. Menzel, A. 2020. ClimateEU, scale-free climate normals, historical time series, and future projections for Europe. Scientific Data 7: 428. https://sites.ualberta.ca/~ahamann/data/climateeu.html] (access date, 4 July, 2020).
Matías, L., Gonzalez-Díaz, P., Quero, J. L., Camarero, J. J., Lloret, F., & Jump, A. S. 2016. Role of geographical provenance in the response of silver fir seedlings to experimental warming and drought. Tree Physiology, 36(10), 1236–1246.
Matović, B. 2013. Analyzes and projections of the impact of climate change using the regional climate model on the future distribution and growth of the main tree species in Montenegro – Final report [in Serbian], Ref. No. UNDP MNE 13-026: 1-45.
Matović, B., Stojanović D. B., Kesić L., Stjepanović S. 2018. Impact of climate on growth and vitality of Norway spruce at Kopaonik Mountain [in Serbian], Topola No 201/202 (2018) str. 99-116.
Mátyás, C., Berki, I., Bidló, A., Csóka, G., Czimber, K., Führer, E., Gálos, B., Gribovszki, Z., Illés G., Hirka, A., Somogyi, Z. 2018. Sustainability of Forest Cover under Climate Change on the Temperate-Continental Xeric Limits. Forests, 9(8), 489: 1–32.
Mellert, K. H., Ewald, J., Hornstein, D., Dorado-Liñán, I., Jantsch, M., Taeger, S., Zang, C., Menzel, A., Kölling, C. 2015. Climatic marginality: a new metric for the susceptibility of tree species to warming exemplified by Fagus sylvatica (L.) and Ellenberg’s quotient. European Journal of Forest Research, 135(1): 137–152.
Mikac, S., Žmegač, A., Trlin, D., Paulić, V., Oršanić, M., & Anić, I. 2018. Drought-induced shift in tree response to climate in floodplain forests of Southeastern Europe. Scientific Reports, 8(1): 1–12.
Mina, M., Bugmann, H., Klopcic, M., & Cailleret, M. 2015. Accurate modeling of harvesting is key for projecting future forest dynamics: a case study in the Slovenian mountains. Regional Environmental Change, 17(1), 49–64.
Misi, D., Puchałka, R., Pearson, C., Robertson, I., & Koprowski, M. 2019. Differences in the Climate-Growth Relationship of Scots Pine: A Case Study from Poland and Hungary. Forests, 10(3), 243.
Móricz, N., Garamszegi, B., Rasztovits, E., Bidló, A., Horváth, A., Jagicza, A.,Illés, G., Vekerdy, Z., Somogyi Z., Gálos, B. 2018. Recent Drought-Induced Vitality Decline of Black Pine (Pinus nigra Arn.) in South-West Hungary-Is This Drought-Resistant Species under Threat by Climate Change?, Forests, 9(7), 414, 1–20.
Móricz, N., Rasztovits, E., Gálos, B., Berki, I., Eredics, A., & Loibl, W. 2013. Modelling the Potential Distribution of Three Climate Zonal Tree Species for Present and Future Climate in Hungary. Acta Silvatica et Lignaria Hungarica, 9(1): 85–96.
Nedealcov, M., Donica, A. 2019. Climate Limitations of The Beech (Fagus sylvatica) Within the Republic of Moldova Territory, IJEES, Vol. 9 (3): 461–468.
Neuner, S., Albrecht, A., Cullmann, D., Engels, F., Griess, V. C., Hahn, W. A., Härtl F., Kölling C., Staupendahl K., Knoke, T. 2014. Survival of Norway spruce remains higher in mixed stands under a dryer and warmer climate. Global Change Biology, 21(2), 935–946.
Noce, S., Collalti, A., & Santini, M. 2017. Likelihood of changes in forest species suitability, distribution, and diversity under future climate: The case of Southern Europe. Ecology and Evolution, 7(22), 9358–9375.
Panayotov, M. P., Zafirov, N., & Cherubini, P. 2012. Fingerprints of extreme climate events in Pinus sylvestris tree rings from Bulgaria. Trees, 27(1), 211–227.
Paul, C., Brandl, S., Friedrich, S., Falk, W., Härtl, F., & Knoke, T. 2019. Climate change and mixed forests: how do altered survival probabilities impact economically desirable species proportions of Norway spruce and European beech? Annals of Forest Science, 76(1).
Pavlović, L., Stojanović, D., Mladenović, E., Lakićević, M., & Orlović, S. 2019. Potential Elevation Shift of the European Beech Stands (Fagus sylvatica L.) in Serbia. Frontiers in Plant Science, 10: 1–8.
Pecchi, M., Marchi, M., Moriondo, M., Forzieri, G., Ammoniaci, M., Bernetti, I., Bindi, M. and Chirici, G. 2020. Potential Impact of Climate Change on the Forest Coverage and the Spatial Distribution of 19 Key Forest Tree Species in Italy under RCP4.5 IPCC Trajectory for 2050s, Forests 2020, 11, 934.
Pierce, D., 2019. ncdf4: Interface to Unidata netCDF (Version 4 or Earlier) Format Data Files. R package version 1.17.
R Core Team, 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rasztovits, E. 2011. Modelling the future distribution of beech at low-elevation xeric limits - comparison of empirical and stochastic models, Dissertation, University of West Hungary, Faculty of Forestry, Sopron, http://doktori.nyme.hu/361/1/disszertacio.pdf (access date, 21 March, 2020).
Republic Hydrometeorological Service of the Republic of Serbia, 2020. Climatology: 30 years averages. http://www.hidmet.gov.rs/eng/meteorologija/klimatologija_srednjaci.php (access date, 8 September, 2020).
Robin, X., Turck, N., Hainard, A., Tiberti, N., Lisacek, F., Sanchez, J., Müller, M. 2020. pROC: Display and Analyze ROC Curves. R package version 1.16.1.
Robson, T. M., Sánchez-Gómez, D., Cano, F. J., & Aranda, I. 2012. Variation in functional leaf traits among beech provenances during a Spanish summer reflects the differences in their origin. Tree Genetics & Genomes, 8(5), 1111–1121.
Rose, L., Leuschner, C., Köckemann, B., & Buschmann, H. 2009. Are marginal beech (Fagus sylvatica L.) provenances a source for drought tolerant ecotypes? European Journal of Forest Research, 128(4), 335–343.
Ruosch, M., Spahni, R., Joos, F., Henne, P. D., van der Knaap, W. O., & Tinner, W. 2016. Past and future evolution of Abies alba forests in Europe – comparison of a dynamic vegetation model with palaeo data and observations. Global Change Biology, 22(2), 727–740.
Rybníček, M., Čermák, P., Kolář, T., & Žid, T. 2012. Growth responses of Norway spruce (Picea abies (L.) Karst.) to the climate in the south-eastern part of the Českomoravská Upland (Czech Republic). Geochronometria, 39(2): 149–157.
Salamon-albert, É., Lőrincz, P., Pauler, G., Bartha, D., & Horváth, F. 2016. Drought Stress Distribution Responses of Continental Beech Forests at their Xeric Edge in Central Europe. Forests, 7(12), 298: 1–16.
Saltré, F., Duputié, A., Gaucherel, C., & Chuine, I. 2014. How climate, migration ability and habitat fragmentation affect the projected future distribution of European beech. Global Change Biology, 21(2), 897–910.
Schwarz, A. J., Bauhus J. 2019. Benefits of mixtures on growth performance of Silver fir (Abies alba) and European beech (Fagus sylvatica) increase with tree size without reducing drought tolerance, Frontiers in Forests and Global Change, 2(79).
Sidor, C. G., Camarero, J. J., Popa, I., Badea, O., Apostol, E. N., & Vlad, R. 2019. Forest vulnerability to extreme climatic events in Romanian Scots pine forests. Science of The Total Environment, 678: 721–727.
Spathelf, P., van der Maaten, E., van der Maaten-Theunissen, M., Campioli, M., & Dobrowolska, D. 2013. Climate change impacts in European forests: the expert views of local observers. Annals of Forest Science, 71(2): 131–137.
Štajner, D., Popović, B. M., Orlović, S., Ždero Pavlović, R., Blagojević, B. 2017. European Beech (Fagus sylvatica L.) from Serbian Mountains – Capacity to Resist Ecological and Oxidative Stress. Baltic Forestry, 23(2): 374-383.
State Enterprise (SE) “Srbijašume”, Main Forest Tree Species Managed by SE “Srbijašume” [in Serbian]. https://srbijasume.rs/gazdovanje-sumama/sumski-fond/glavne-vrste-sumskog-drveca/ (access date, 26 January, 2021).
State Enterprise (SE) “Vojvodinašume”, Forests, Main tree varieties [in Serbian]. https://www.vojvodinasume.rs/sume/glavne-vrste-drveca/ (access date, 26 January, 2021).
Stjepanović, S., Matović, B., Stojanović. B. D., Levanič, T., Orlović, S., Kesić, L., Pavlović, L. 2017. Influence of Dry Years on Width of Tree Rings оf European Beech (Fagus sylvatica L.) in the Western Republic оf Srpska [in Serbian], Topola No 199/200: 141–152.
Stojanović, D. B, Matović, B., Orlović, S., Kržič, A., Trudić, B., Galić, Z., Stojnić, S., Pekeč, S. 2014. Future of the Main Important Forest Tree Species in Serbia from the Climate Change Perspective. SEEFOR 5 (2): 117–124.
Stojanović, D. B., Kržič, A., Matović, B., Orlović, S., Duputie, A., Djurdjević, V., Galić, Z., Stojnić, S. 2013. Prediction of the European beech (Fagus sylvatica L.) xeric limit using a regional climate model: An example from southeast Europe. Agricultural and Forest Meteorology, 176: 94–103.
Stojanović, D. B., Levanič, T., Matović, B. 2015b. Correlation between different climate variables and indices and growth of Turkey oak (Quercus cerris L.) [in Serbian], Topola/Poplar No 195/196: 23-29.
Stojanović, D. B., Levanič, T., Matović, B., & Orlović, S. 2015a. Growth decrease and mortality of oak floodplain forests as a response to change of water regime and climate. Eur J Forest Res, 134(3): 555–567.
Stojanović, Lj., Krstić, M. 2009. Thinning in artificially established stands of Austrian pine and Scots pine in Serbia [in Serbian], Šumarstvo br. 1-2, Beograd, 1–21.
Swets J. (1988) Measuring the accuracy of diagnostic systems. Science 240, 1285- 1293.
Taeger, S., Zang, C., Liesebach, M., Schneck, V., & Menzel, A. 2013. Impact of climate and drought events on the growth of Scots pine (Pinus sylvestris L.) provenances. Forest Ecology and Management, 307, 30–42.
Taleshi, H., Jalali, S. G., Alavi, S. J., Hosseini, S. M., Naimi, B., & Zimmermann, N. E. 2019. Climate change impacts on the distribution and diversity of major tree species in the temperate forests of Northern Iran. Regional Environmental Change, 19:2711–2728.
Thurm, E. A., Hernandez, L., Baltensweiler, A., Ayan, S., Rasztovits, E., Bielak, K., Mladenov Zlatanov, T., Hladnik, D., Balic, B., Freudenschuss, A., Büchsenmeister, R., Falk, W. (2018). Alternative tree species under climate warming in managed European forests. Forest Ecology and Management, 430, 485–497.
Tikvić, I., Seletković, Z., Ugarković, D., Posavec S., Španjol, Ž. 2008. Dieback of Silver Fir (Abies alba Mill.) on Northern Velebit (Croatia), Periodicum biologorum, 110(2): 137–143.
Tomić, Z., Rakonjac Lj, Isajev, V. 2011. Monograph: The Selection of Species for Reforestation and Amelioration in Central Serbia [in Serbian], Institute of Forestry, Belgrade.
Trombik, J., Hlásny T., Dobor, L., Barcza, Z. 2013. Climatic Exposure of Forests In The Carpathians: Exposure Maps And Anticipated Development, International Scientific Conference for PhD Students March 19-20 2013: 1–9, Győr, Hungary.
Vlăduţ, A., Nikolova, N., Licurici, M. 2017. Influence of Climatic Conditions On The Territorial Distribution Of The Main Vegetation Zones Within Oltenia Region, Romania, Muzeul Olteniei Craiova. Oltenia. Studiişicomunicări. Ştiinţele Naturii. Tom. 33(1): 154–164.
Walentowski H., Falk W., Mette T., Kunz J., Bräuning A., Meinardus C., Zang Ch., Sutcliffe L., Leuschner Ch., 2017. Assessing future suitability of tree species under climate change by multiple methods: a case study in southern Germany. Ann. For. Res. 60(1): _–_



