Abstract
Deadwood is one of the important components of forest ecosystems since it is considered as an indicator of resource availability and biodiversity, an important carbon stock, and habitat for insect and fungal species. Removal of dead trees from forests may result in the elimination of many species, and also reduce the number and density of species. Trojan fir (Abies nordmanniana subsp. equi-trojani) is known to be rich in species composition; however, our knowledge on the amount and type of deadwood (i.e. standing or lying) in these forests is limited. Thus, in this study, deadwood quantity and type in Trojan fir forests were determined. In addition, less is known about the factors that influence the amount of deadwood volume; therefore, the effects of stand type, elevation (m), aspect, quadratic mean diameter (QMD) (cm), basal area (BA) (m2 ha-1) and stand volume (m3 ha-1) on deadwood volume were also observed. Stand type, QMD and BA significantly affected the volume of deadwood in Trojan fir forests while stand volume, elevation and aspect had no influence on the deadwood volume. In comparison to the values in the literature, the amount of deadwood observed seems to be acceptable across all stand types. Given the initial results from this study, it can be recommended that large scale deadwood assessments are needed in Turkey, and it can be combined with the current inventory system.
1. Introduction
Deadwood is a general term used to define dead or dying trees that are fallen on the ground (logs) or standing (snags) in forest ecosystems (Mark et al. 2006). Deadwood is considered vital for the conservation and sustainability of biodiversity in forests (Samuelsson et al. 2005), and it is also important source for many microorganisms to survive (Junninen et al. 2006; O´dor et al. 2006; Ulikzka and Angelstam 2000; Davies et al. 2008; DeMaynadier and Hunter 1995; Martin and Eadie 1999; Sullivan and Sullivan 2001). Deadwood also plays a key role in the growth, survival and establishment of understory seedlings by increasing water holding capacity of the soil, and enhancing seedbed for germination (Christensen et al. 2005). In addition, since forests are one of the biggest carbon (C) pools on the earth and they are crucial for carbon sequestration, decomposition of deadwood in forests plays a key role in its C cycle (Weggler et al. 2012; Köster et al. 2015). Therefore, more detailed data on deadwood is needed to study its impacts on biodiversity, C stocks, regeneration, etc.
Recently, the role of deadwoods in sustainability of biodiversity has been studied in different countries (Christensen et al. 2005; Davies et al. 2008). It has been suggested that higher amount of deadwood usually results in higher resource availability, and consequently, higher number and density of species (Bassler and Müller 2010; Müller and Bütler 2010). Recently, the influence of deadwood on forest establishment and biodiversity has gained recognition among scientists (Hunter 1990; Jonsson et al. 2005; Atıcı et al. 2008). These studies have been usually focused on the composition and dynamics of dead trees and their interactions with animals, plants, insects and fungi (Harmon et al. 1986; Fridman and Walheim 2000; Siitonen 2001; Fraver et al. 2002; Laiho and Prescott 2004; Steephenes and Moghaddas 2005; Çolak et al. 2009; Atıcı et al. 2008). Due to increasing interest on the sustainable forestry and conservation of biodiversity, there should be more effort to increase the amount of deadwoods in managed forests (Kirby et al. 1998; Christensen et al. 2005; Marage and Lemperiere 2005). The importance of deadwoods for biodiversity has been globally accepted, but guidelines on this aspect of forest management have been limited (Müller and Bütler 2010). Inventory of deadwood in varying stand structures and determination of factors that influence the amount of deadwood seem to be necessary for future planning, conservation of biodiversity, and carbon cycle.
Trojan fir (Abies nordmanniana subsp. equi-trojani) is a shade tolerant tree species demonstrating uneven-aged structures in vast areas in Turkey (Güner et al., 2012). This species mostly occurs at elevations ranging from 100 to 1800 m. Trojan fir can be found as pure and mixed stands in Turkey, and covers an area of approximately 670.000 hectare which is 3.1% of the total forested land in the country (General Directorate of Forestry 2014). This species is considered as one of the most important tree species of Turkey due to its high-quality timber as well as its aesthetic values in the landscapes (Sevik 2012). Trojan fir can survive under canopy for a prolonged time without losing its growth potential (Saraçoğlu 1988).
Forests in Turkey are considered rich in biodiversity; thus, there is a need for new silvicultural prescriptions that ensures biodiversity conservation while producing high quality timber (Çolak et al. 2009; Kurt et al. 2015). Although there has been an increasing interest on the retention of deadwoods worldwide, current studies on the deadwood in Turkey has been limited (Ulbrichova et al. 2016). To our knowledge, there has not been enough research on the investigation of the amount and type of deadwood in Trojan fir forests. In addition, it has been stated that amount of deadwood depends on several factors including tree species, stand type, stand age, site index, aspect, elevation, forest management activities, wood quality and the speed of decomposition (Simila et al. 2003; Heilmann-Clausen and Christensen 2004; Fischer 2008). Thus, in this study, our main objectives were to 1) determine the deadwood quantity and type in Trojan fir forests, and 2) observe the influences of stand type, elevation (m), aspect, quadratic mean diameter (QMD) (cm), basal area (BA) (m2 ha-1) and stand volume (m3 ha-1) on deadwood volume. With the obtained data, we aim to enhance our knowledge about deadwood that is important for the biodiversity of Trojan fir forests in Turkey. This study will also help make better decisions in silvicultural prescriptions, and encourage future deadwood studies within the region.
2. Materials and methods
2.1 Study area
This study was conducted in the Küre Forest Planning Directorate (KFPD) located in Kastamonu, Turkey. KFPD covers an area of approximately 142.000 hectare; 69% of the total area is forested while agricultural, pasture and residential areas cover 25, 4 and 2 % of the total area, respectively. The forests in Küre are dominated by Trojan fir trees, and mostly pure with varying stand structures. These forests have been commonly managed using selection silviculture. The area is located within the Euro-Siberian fito geographic region (Çolak et al. 2009), which is represented by the forests that composed of broad-leaf and needle-leaf trees. Research area shows the typical characteristic of terrestrial Black Sea climate with cold winters and rainy summers. Average annual precipitation is approximately 900 mm. Within the study area, brown forest soil types are common. Scots pine (Pinus sylvestris L.), European aspen (Populus termula L.), European hornbeam (Carpinus betulus L.), bitter willow (Salix elaeagnos) and Oriental beech (Fagus orientalis) are other common tree species in the study. Salvia glutinosa L., Verbascum eriocarpum, Primula vulgaris and Rosa dumalis are some of the understoy plants within the study area.
2.2 Plot design
In Turkey, uneven-aged Trojan fir stands are divided into four categories (GA, GB, GC and GD stands) based on the stand structures (Eraslan 1982). GA stands are old uneven-aged fir stands, close to optimum (i.e. sustainable diameter distribution-that is to say number of trees in each diameter class) and contain mostly large diameter trees. GB stands are young uneven-aged stands, and they mostly consist of small size fir trees. Compared to the optimum, GC stands contain mostly middle to large diameter trees, and they usually present even-aged structures with one canopy tier. GD stands lack of middle size trees, and contain mostly small and large diameter trees with uneven-aged structure (Eraslan 1982). In the study area, potential stands with different stand structures (i.e. GA, GB, GC and GD) were determined from managed Trojan fir forests within the five Forest Planning Units of KFPD in 2012. Then, 10 study plots were randomly installed within each stand type (i.e., 4x10=40 plots in total). Study plots were in size of 0.25 ha (50x50 m) as suggested by Atici et al. (2008) (Figure 1).
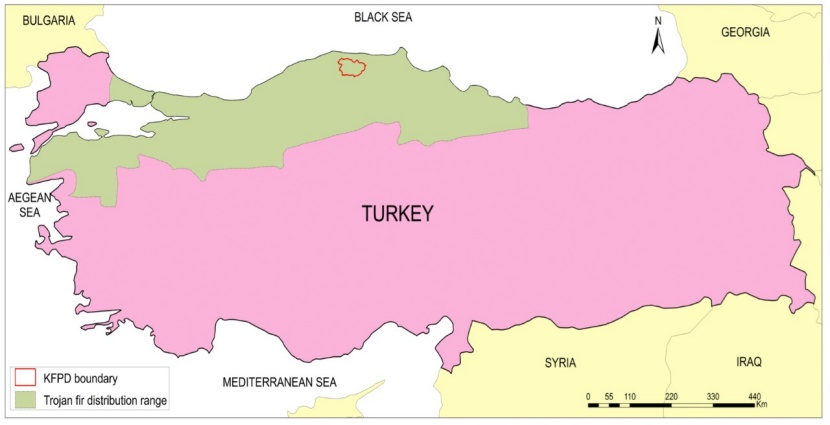
Figure 1: Trojan fir distribution range and the location of the research area (Bizimbitkiler, 2017)
2. 3 Measurements and analysis
Elevation and aspect of each study plot were recorded. In addition, QMD and stand BA were estimated following the measurements of diameters at breast height (dbh) of all standing live trees with a dbh of 5 cm and greater within each study plot. Tree height was measured, number of trees per ha was recorded and stand density index (SDI) (Reineke 1933) was calculated for each study plot.
In practice, deadwoods are usually classified as standing deadwood (SDW) (i.e. snags) or fallen deadwood (FDW) (i.e. logs) (Mark et al. 2006). SDW was divided into 2 categories; SDW1 and SDW2 (McComb and Lindenmayer 1999; Mark et al. 2006). SDW1 refers to the standing dead trees with a dried up and intact top while SDW2 refers to the standing dead trees with a sloughed off bark and broken top (McComb and Lindenmayer 1999; Mark et al. 2006). FDW was also divided into 2 categories; FDW1 and FDW2 (McComb and Lindenmayer 1999; Mark et al. 2006). FDW1 represents to the recently fallen dead trees while SDW2 refers to the rotten fallen dead trees (McComb and Lindenmayer 1999; Mark et al. 2006). For the coarse deadwoods, a tree with a diameter of 10 cm and above is used as the standard size in practice and scientific studies. (Swanson et al. 1976; Ranius et al. 2003; Norden et al. 2004; Webster and Jenkins 2005; Lipan et al. 2008; Atıcı et al. 2008; Beets et al. 2008; Çolak et al. 2009). Deadwood with a diameter smaller than 10 cm is not as important as coarse deadwoods (Coomes et al. 2002; Norden et al. 2004; Beets et al. 2008; Lipan et al. 2008; Çolak et al. 2011), therefore, deadwoods smaller than 10 cm in diameter was not included in the analysis in this study (Comes et al. 2002; Norden et al. 2004; Beets et al. 2008).
In order to calculate volume of living wood (LW) (m3 ha-1) and SDW1 (m3 ha-1) in each plot, height (h) and dbh of trees were recorded. The volume tables for Trojan fir created by Miraboglu (1955) were utilized when estimating LW and SDW1. Volume of SDW2 trees was estimated as the sum of the standing part and fallen parts of same trees on the ground using the equation 1 (Husch et al. 1993).
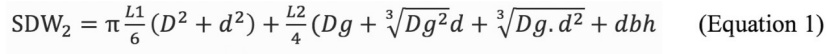
where L1 is length of upper part, L2 is length of lower part, d is the diameter of the top of the lower part, D is the diameter of the end of the upper part, Dg is diameter at the ground level. FDW was calculated using the equation 2 (Patterson et al. 2007). It has been stated that equation 2 is insensitive to the changes in species and site (Çolak et al. 2009).
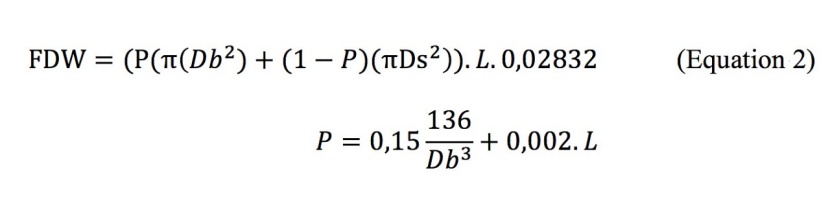
where Ds is the diameter at the small end, Db is the butt end diameter, and L is the length of the log. When there was a stem deformation, the diameter at stump height (dsh) (d0.3) was measured, and then dbh was calculated using the relationships between dbh and dsh. Total deadwood (TDW) (m3 ha-1) was calculated as the sum of SDW and FDW. In addition, TDW/LW ratio (%) was calculated for each plot.
Influence of stand type, elevation, aspect, QMD, BA and stand volume on deadwood volume were determined using multiple linear regression with the R programming language (R Development Core Team 2010). A mixed-effect model that incorporated stand type, elevation, aspect, QMD, BA and stand volume as a fixed-effect and plots as random effects was used to calculate the significance of the values. Multiple comparisons of means of stand types were done using the Tukey’s method in the R programming language (R Development Core Team 2010).
3. Results
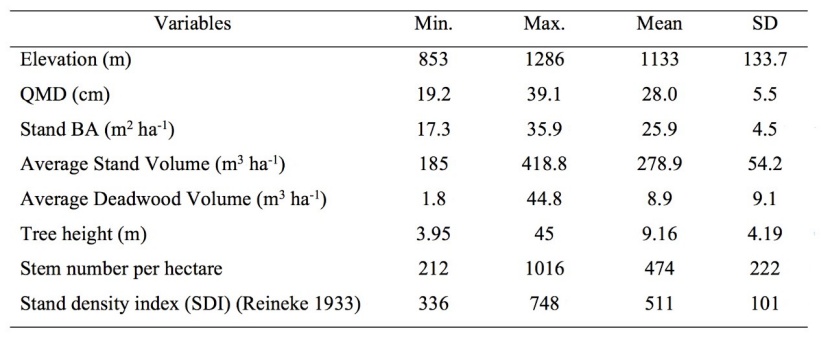
Table 1: Descriptive statistics of parameter used for their influence on the total deadwood volume.
Descriptive statistics of parameters including elevation, QMD, BA, total stand volume, total deadwood volume, tree height, stem number per ha and SDI were given in Table 1. Average deadwood volume was 8.9 m3 ha-1 across all study plots (Table 1). Given the total acreage of the study area, in average, approximately 1,264,000 m3 of deadwood seems to be present within the study area.
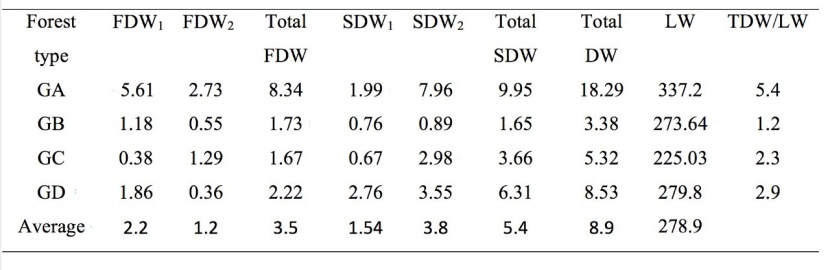
Table 2: Quantitative description of deadwood classes (m3 ha-1) and LW (m3 ha-1), and TDW/LW ratio (%) for each forest type. FDW is fallen deadwood, SDW is standing deadwood, LW is living wood, and TDW is total deadwood.
Table 2 shows all deadwood classes including SDW1, SDW2, FDW1 and FDW2, as well as TDW, LW and TDW/LW ratios in different stand types (i.e. GA, GB, GC and GD) of Trojan fir forests. The amount of FDW ranged from 1.67 to 8.34 m3 ha-1 (average 3.49 m3 ha-1) across all stand types; largest FDW was observed in GA followed by GD and GC, while lowest FDW was in GB (Table 1). SDW and TDW values demonstrated the same pattern; we observed the largest SDW (9.95 m3 ha-1) and TDW (18.29 m3 ha-1) in GA followed by GD and GC, and the lowest in GB (1.65 and 3.38 m3 ha-1, respectively) (Table 1). As for LW, the highest LW value (337.2 m3 ha-1) was monitored in GA followed by GD and GB, and the lowest (225.3 m3 ha-1) in GC (Table 1). TDW/LW ratio ranged from 0.01 to 0.05 (average 0.03) across all stand types; it was biggest in GA followed by GD, GC and GB.

Table 3: Average deadwood volume (m3 ha-1) for each stand type based on deadwood diameters
Since some studies highlighted the importance of deadwood diameter (Bouget et al. 2014), we divided deadwood volume into 2 classes; small deadwood and large deadwood as suggested by Bouget et al. (2014). Table 3 gives the average deadwood volume for each stand type based on deadwood diameters.
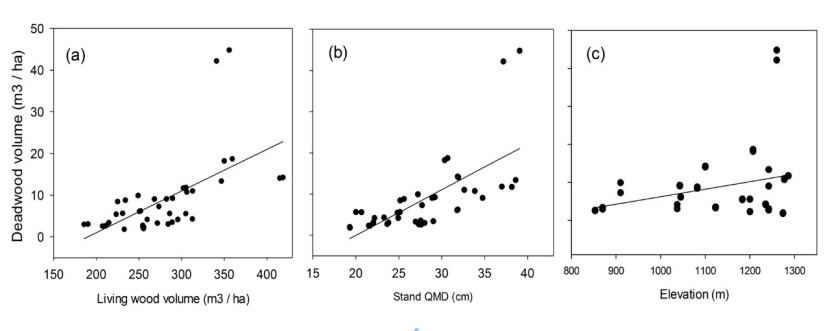
Figure 2: Relations between deadwood volume and (a) living wood volume, (b) stand QMD, and (c) elevation.
When the total volume of deadwood and living wood were compared across all stand types, it was found that there was a statistically significant relationship between the deadwood volume and living wood volume (p=0.012). However, it can be seen on the Figure 2a that deadwood volume did not significantly change (ranging from 1 to 10 m3 ha-1) with increasing stand volume until the stand volume reaches approximately 300 m3 ha-1 (Figure 2a). In denser stands, total deadwood increased with increasing stand volume (Figure 2a). In addition, we also observed a statistically significant relationship between QMD of stand and total volume of deadwood (p=0.026) (Figure 2b), between BA and total volume of deadwood (p=0.035), and between SDI and total volume of deadwood (p<0.05) in Trojan fir forests. There was no significant influence of either elevation or aspect on the total volume of deadwood (p=0.204 and p=0.214 for elevation and aspect, respectively) (Figure 2c). The two plots with fairly high deadwood volume (Figure 2) were from GA plots which contain mostly large diameter trees. Because average dbh and height of dead trees were substantially high (46.5 cm and 20 m, respectively) within these GA plots, they seemed to be away from the distribution. It is likely that these plots may simply be within the inherent range of variability in GA stands, thus, they were retained in the analysis.
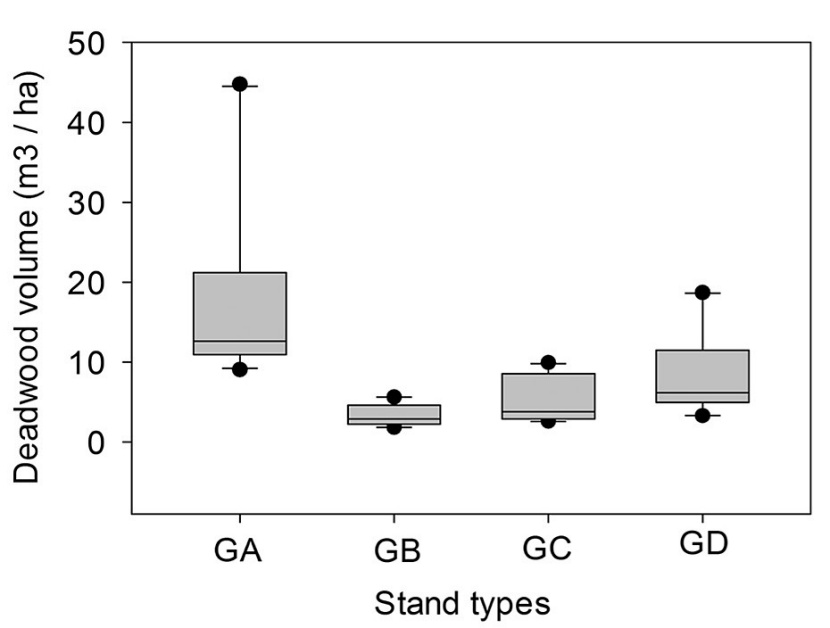
Figure 3: Relation between deadwood volume and stand types.
Stand type has a significant influence on the total volume of deadwood in Trojan fir forests (p<0.0001) (Figure 3). In addition, when we observed the influence of same parameters (i.e. stand type, elevation, aspect, QMD, BA and stand volume) on the volume of given deadwood categories (i.e. FDW1, FDW2, SDW1 and SDW2), we determined that QMD significantly affected the volume of FDW1, SDW1 and SDW2. Tukey multiple comparisons of means showed that there were significant relationships between GA and GB (p=0.0001), between GA and GC (p=0.0004), and between GA and GD stands (p=0.009). No significant relationships were observed between GB and GC (p=0.901), between GB and GD (p=0.285), and between GC and GD stands (p=0.673).
4. Discussion
This study aimed to determine whether present conditions in Trojan fir forests provide sufficient amount of deadwood. Utschik (1991) and Scherzinger (1996) stated that the volume of deadwood should be between 1-3 m3 ha-1 in managed forests for biodiversity. Studies conducted in Finland, Sweden, Germany, France, Belgium, and Switzerland have showed that the average TDW volume in managed forests is less than 10 m3 ha-1 (Christensen et al. 2005). Our average TDW value (8.88 m3 ha-1) seems to be relatively high, and it can be considered satisfactory given the values in the literature (Ammer 1991; Utschik 1991). The amount of deadwood in unmanaged forests is usually between 5 and 30% (Fridman and Walheim 2000). It has been suggested that the minimum amount of deadwood in a managed forest changes depending on the site, but it should be around 1-2% of the whole yield (Ammer 1991). An average of 3% of the TDW/LW ratio observed in this research also indicates that our TDW/LW ratio was mostly within the suggested range, and relatively higher than the recommended value of 1-2% (Table 2). In addition, Albrecht (1991) stated that there should be 2-5 dead trees per hectare to maintain sustainable biodiversity in managed forests. In this study, we observed a significantly higher number of dead trees per hectare (27 trees ha-1 in total, and 16 trees ha-1 as snags) than suggested range. It has been recommended that biodiversity level of a forest is associated with the amount of deadwood volume it contains (Bartoli and Geny 2005; Debeljak 2006; Bassler and Müller 2010; Müller and Bütler 2010).
Although biodiversity of a forest is associated with the amount of deadwood volume it contains (Bartoli and Geny 2005; Debeljak 2006; Böhl and Brändli 2007; Bassler and Müller 2010; Müller and Bütler 2010), the amount of varying types of deadwood (i.e. snags or fallen trees) is also essential for specific objectives. For example; Ammer (1991) pointed out the importance of snag volume in a stand, and suggested a deadwood volume of 5-10 m3 ha-1 with at least 50% as snags for bird conservation. In our study, percentage of snags (i.e. standing deadwoods) was 61% of the total deadwood volume, which seemed to be satisfactory for wildlife conservation. In addition, some studies highlighted the importance of deadwood diameter (Bouget et al. 2014). Our deadwood volumes by diameter groups (i.e. small and large deadwood) are consisted with the stand structure. For example, GA stands mostly consists of large trees, thus, we observed most large deadwood (i.e. deadwood larger than 40 cm in diameter) in this stand type. It has been suggested that large deadwood can enhance biodiversity in an ecosystem (Bouget et al. 2014). On the other hand, there was no large deadwood in GB stand type because these stands consist of small diameter fir trees. Given these observations, GA and GD stands can exhibit more diverse conditions compared to GB and GC.
Since Trojan fir forests have been managed using selection silviculture under high residual stand density (Odabaşı et al. 2004), it is possible to observe density-related mortality resulting in high or satisfactory amount of deadwood volume in these forests. This was substantiated by our findings as BA, QMD and SDI significantly influenced the amount of deadwood volume across all stand types. In addition, the significant relationships between the living wood volume and deadwood volume (Figure 2) support the assumption that the amount of deadwood volume can be attributed to the density-related mortality in Trojan fir forests. Atici et al. (2008) stated that stand volume above 100 m3 ha-1 may increase the risk for insect damage in some forest types indicating the indirect effects of stand density on the amount of deadwood volume. Moreover, it has been recommended that silvicultural treatments and harvesting can directly and indirectly impact the quantity, quality, and dynamics of deadwood resources (Müller- Using and Bartsch 2003; Saniga and Schütz 2001). Because selection methods and other types of partial cuts are considered higher risk of damage to residual trees due to more frequent disturbances in these methods (Lamson et al. 1985), they may have affected the amount of deadwood volume in Trojan fir forests as well. Conversely, it is likely that even-aged silvicultural systems may result in less amount of deadwood due to higher amount of removal and lower stand densities in these systems.
Harmon et al. (1986) stated that the amount of deadwood is usually lower in managed forests compared to unmanaged old-growth forests because managed forests usually lack of large size trees. Our findings substantiate this statement. It should be noted that GA stands refers to old uneven-aged fir stands that are close to optimum, and contain mostly large diameter trees. Even though GA stands are also managed, high number of large diameter trees creates relatively higher amount of deadwood volume in these stands. At the other end of the spectrum, GB stands, which mostly contain young and small size trees, had the lowest amount of total deadwood due to the scarcity of large and old trees in GB stands. It has been aimed to increase the amount of deadwood in managed forests (Christensen et al. 2005; Marage and Lemperiere 2005), but present silvicultural prescriptions seem to be favorable to obtain adequate amounts of deadwood in managed GA stands in Turkey. Our findings also recommend prescriptions that aim to keep some large and old fir trees as reserve during regeneration process so that young stands (such as GB stands) can contain sufficient number of dead trees.
5. Conclusions
With comprehensive researches, introduction of new silvicultural prescriptions and regulations regarding deadwood volume seems to be vital in forest of Turkey. The preliminary data presented in this study aim to broaden our understanding of how stand parameters such as stand type, BA and QMD influence the amount of deadwood volume in Trojan fir forests in Turkey. In addition, the current data also help for future silvicultural prescriptions that aim to ensure sustainable biodiversity conservation while producing high quality timber. Given the values in the literature, our data may suggest that the Trojan fir forests already exceed the recommended deadwood volume. But, future monitoring is required to determine how and when the amount of deadwood volume reaches the acceptable levels in young Trojan fir stands. In addition, further studies on the assessment of deadwood in unmanaged Trojan fir stands, as well as its comparison with managed stands, are also recommended. Finally, managers need to monitor the effectiveness of their management plans so that they can adjust prescriptions using a responsive management approach.
Acknowledgments
This research has been supported by Kastamonu University Scientific Research Projects Coordination Department. Project Number: KÜBAP 01/2013-53.
References
Albrecht, L. (1991). Die Bedeutung des toten Holzes im Wald. Forstwissenschaftliches Centralblatt 110: 106-113.
Ammer, U. (1991). Konsequenzen aus den Ergebnissen der Totholzforschung für die forstliche Praxis. Orstwissenschaftliches Centralblatt 110: 149-157.
Atici, E., Çolak, AH., Rotherham, ID. (2008). Coarse Dead Wood Volume of Managed Oriental Beech (Fagus orientalis Lipsky.) Stands in Turkey. Investigación Agraria: Sistemas y Recursos Forestales 17(3): 216-227.
Bartoli, M., Geny, B. (2005). Once upon a time... dead wood in French forests. Revue Forestiere Francaise 57(5): 443-456.
Bässler, C., Müller, J. (2010). Importance of natural disturbance for recovery of the rare polypore Antrodiella citrinella Niemela & Ryvarden. Fungal Biology 114:129–133.
Beets, PN., Hood, IA., Kimberley, MO., Oliver, GR., Pearce, SH., Gardner, JF. (2008). Coarse woody debris decay rates for seven indigenous tree species in the central North Island of New Zealand. Forest Ecology and Management 256 (4): 548-557.
Bizimbitkiler. (2017). Kazdağı göknarı yayılışı. Available at http://www.
bizimbitkiler.org.tr/v2/sonuc.php?i=VmxMFlWbFdWWGhWYmxKV1YwZFNUMVpzV21GaU1WWnhVbTVrV2xadGVIcFhhMUpQVkcxS1NHVkdhRmhXUlRWMldWVmFXbVF4WkhOalJtUlhaV3hhVFZkV1ZtRlhiVlowVW10c1ZXSklRazlaYkZWM1RWWmFWbGRyV2xCV2EwcFRWVVpSZDFCUlBUMD0=
Bouget, C., Larrieu, L., Brin, A. (2014). Key features for saproxylic beetle diversity derived from rapid habitat assessment in temperate forests. Ecological Indicators 36: 656-664.
Böhl, J., Brändli, UB. (2007). Deadwood volume assessment in the third Swiss National Forest Inventory: methods and first results. European Journal of Forest Research 126(3): 449-457.
Christensen, M., Hahn, K., Mountford, EP., Ódor, P., Standovár, T., Rozenbergar, D., Diaci, J., Wijdeven, S., Meyer, P., Winter, S., Vrska, T. (2005). Dead wood in European beech (Fagus sylvatica) forest reserves. Forest Ecology and Management 210 (1-3): 267-282.
Coomes, DA., Allen, RB., Scott, NA., Goulding, C., Beets, P. (2002). Designing systems to monitor carbon stocks in forests and shrublands. Forest Ecology and Management 164: 89-108.
Çolak, AH., Tokcan, M., Rotherham, ID., Atici, E. (2009). The amount of coarse dead wood and associated decay rates in forest reserves and managed forests, northwest Turkey. Investigación Agraria: Sistemas y Recursos Forestales 18 (3): 350-359.
Çolak, AH., Tokcan, M., Kirca, S. (2011). Dead Wood; Unseen Life on Dead. The Western Blacksea Forestry Research Institute- Bolu Ministry of Environment and Forestry-Turkey Various Publications Series Nr. 5
DeMaynadier, PG., Hunter, ML. (1995). The relationship between forest management and amphibian ecology: a review of the North American literature. Environmental Reviewes 3: 230–261.
Davies, ZG., Tyler, C., Stewart, GB., Pullin, AS. (2008). Are current management recommendations for conserving saproxylic invertebrates effective? Systematic Review 17: 209–234.
Debeljak, M. (2006). Coarse woody debris in virgin and managed forest. Ecological Indicators 6: 733-742.
Eraslan, İ. (1982). Orman Amenajmanı Ders Kitabı, İstanbul Üniversitesi Orman Fakültesi Yayınları, İstanbul. (In: Turkish)
Fischer, G. (2008). Aktiv für Totholz im Wald. Anregungen für Forstleute und Landwirte. Österreichische Bundesforste AG, Kompetenzfeld Naturund Umweltschutz, 3002, Purkersdorf.
Fraver, S., Wagner, RG., Day, M. (2002). Dynamics of coarse woody debris following gap harvesting in Acadian forest of central Maine, USA. Canadian Journal of Forest Research 32: 2094-2105.
Fridman, J., Walheim, M. (2000). Amount, structure and dynamics of dead wood on managed forestland in Sweden. Forest Ecology and Management 131: 23-36.
General Directorate of Forestry (2014). Forest Atlas. Publications of General Directorate of Forestry. 116 p.
Güner, A., Aslan, S., Ekim, T., Vural M., Babaç, M. T. (2012). Türkiye Bitkileri Listesi (Damarlı Bitkiler). Nezahat Gökyiğit Botanik Bahçesi ve Flora Araştırmaları Derneği Yayını. İstanbul. ISSN: 978-605-60425-7-7.
Harmon, ME., Franklin, JF., Swanson, FJ., Sollins, P., Gregory, SV., Lattin, JD., Anderson, NH., Cline, SP., Aumen, NG., Sedell, JR., Lienkaemper, GW., Cromack, KJR., Cummins, KW. (1986). Ecology of coarse woody debris in temperate ecosystem. In: Advances in Ecological Research, Academic Press, New York, 15: 133-302.
Heilmann-Clausen, J., Christensen, M. (2004). Does size matter? On the importance of various dead wood fractions for fungal diversity in Danish beech forests. Forest Ecology and Management 201: 105–117.
Hunter, ML. (1990). Wildlife, forests and forestry: Principles of managing forests for biological diversity. Engelwood Cliffs, Prentice-Hall, New York
Husch, BC., Miller, I., Beers, TW. (1993). Forest mensuration. Third edition. Krieger Publication Co., Malabar, Florida, USA. 402 p.
Jonsson, BG., Kruys, N., Ranius, T. (2005). Ecology of species living on dead wood – Lessons for dead wood management. Silva Fennica 39: 289-309.
Junninen, K., Simila, M., Kouki, J., Kotiranta, H. (2006). Assemblages of wood-inhabiting fungi along the gradients of succession and naturalness in boreal pine-dominated forests in Fennoscandia. Ecography 29: 75–83
Karjalainen, L., Kuuluvainen, T. (2002). Amount and diversity of coarse woody debris within a boreal forest landscape dominated by Pinus sylvestris in Vienasalo wilderness, eastern Fennoscandia. Silva Fennica 36: 147-167.
Kirby, KJ., Reid, CM., Thomas, RC., Goldsmith, FB. (1998). Preliminary estimates of fallen dead wood and standing dead trees in managed and unmanaged forests in Britain. Journal of Applied Ecology 35: 148-155.
Koster, K., Metslaid, M., Engelhart, J., Koster, E. (2015). Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests. Forest Ecology and Management 354: 35-42.
Kurt, Y., Frampton, J., Isik, F., Landgren, C., Chastagner, G. (2016). Variation in needle and cone characteristics and seed germination ability of Abies bornmuelleriana and Abies equi-trojani populations from Turkey. Turkish Journal of Agriculture and Forestry 40 (2): 169-176.
Lamson, NI., Smith, HC., Miller, GW. (1985). Logging Damage Using an Individual-Tree Selection Practice in Appalachian Hardwood Stands. North Journal of Applied Forestry 2: 117-120.
Laiho, R., Prescott, CE. (2004). Decay and nutrient dynamics of coarse woody debris in northern coniferous forest: a synthesis. Canadian Journal of Forest Research 34: 763-777.
Lipan, Y., Wenyao, L., Wenzhang, M. (2008). Woody debris stocks in different secondary and primary forests in the subtropical Ailao Mountains Southwest China. Ecological Research 23: 805-812.
Marage, D., Lemperiere, G. (2005). The management of snags: A comparison in managed and unmanaged ancient forests of the Southern French Alps. Annals of Forest Science 62(2): 135-142.
Mark, CV., Malcolm, JR., Smith, SM. (2006). An integrated model for snag and downed woody debris decay class transitions. Forest Ecology and Management 234(1-3): 48-59.
Martin, K., Eadie, JM. (1999). Nest webs: a community-wide approach to the management and conservation of cavity-nesting forest birds. Forest Ecology and Management 115: 243-257.
Mccomb, W., Lindenmayer, D. (1999). Dying, dead, and down trees, In: Hunter Jr M.L. (Ed.) Maintaining Biodiversity in Forest Ecosystems (pp. 335-372). Cambridge University Press, Cambridge.
Miraboglu, M. (1955). Göknarlarda sekil ve hacim arastirmalari. General Directory of Forestry, Ankara. (In:Turkish).
Müller, J., Bütler, R. (2010). A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. European Journal of Forest Research, 129: 981-992.
Müller-Using, S., Bartsch, N. (2003). Dynamics of woody debris in a beech stand (Fagus sylvatica L.) in Solling. Input, causes and decomposition of woody debris. Allgemeine Forst und Jagdzeitung 174(1): 122-130.
Norden, B., Gotmark, F., Tonnberg, M., Ryberg, M. (2004). Dead wood in semi-natural temperate broadleaved woodland: contribution of coarse and fine dead wood, attached dead wood and stumps. Forest Ecology and Management 194(1-3): 235-248.
Odabaşı, T., Calışkan, A., Bozkuş, HF. (2004). “Silvikültür tekniği”. İstanbul Üniversitesi Yayınları. Yayın no: 4459. (In:Turkish).
O ´dor, P., Heilmann-Clausen, J., Christensen, M., Aude, E., Dort v, KW., Piltaver, A., Siller, I., Veerkamp, MT., Standova´r, T., Hees v, AFM., Kosec, J., Matocec, N., Kraigher, H., Grebenc, T. (2006). Diversity of dead wood inhabiting fungi and bryophytes in semi-natural beech forests in Europe. Biological Conservation 131: 58-71.
Patterson, DW., Doruska, PF., Hartley, J., Hurd, M. (2007). Validating the Patterson and Doruska equation for estimating volume of hardwood butt logs. Forest Products Journal 57: 67-70.
Reineke, L. (1933). Perfecting a stand-density index for even-aged forests. Journal of Agricultural Research 46: 627–638.
Saniga, M., Schütz, JP. (2001). Dynamics of changes in dead wood share in selected beech virgin forests in Slovakia within their development cycle. Journal of Forest Science 47: 557-565.
Saraçoğlu, Ö. (1988). Karadeniz Yöresi Göknar Meşcerelerinde Artım ve Büyüme, Doktora Tezi, Orman Genel Müdürlüğü, İstanbul.
Samuelsson, J., Gustafsson, L., Ingelög, T. (1994). Dying and dead trees – a review of their importance for biodiversity. Swedish Environmental Protection Agency, Uppsala, 109 p.
Scherzinger, W. (1996). Naturschutz im Wald, Qualitätsziele einer dynamischen Wald-entwicklung. Praktischer Naturschutz. Verlag Eugen Ulmer, Stuttgart.
Sevık, H. (2012). Variation in seedling morphology of Turkish fir (Abies Trojaniana subsp. bornmulleriana Mattf). African Journal of Biotechnology 11(23): 6389-6395.
Siitonen, J. (2001). Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forest as an example. Ecological Bulletins 49: 11–41.
Simila, M., Kouki, J., Martikainen, P. (2003). Saproxylic beetles in managed and seminatural scots pine forests: quality of dead wood matters. Forest Ecology and Management 174: 365-381.
Stephen, SL., Moghaddas, JJ. (2005). Fuel treatment effects on snag and coarse woody debris in a Sierra Nevada mixed conifer forest. Forest Ecology and Management 214 (1-3): 53-64.
Swanson, FJ., Lienkaemper, GW., Sedell, JR. (1976). History, physical effects and management implications of large organic debris in Westem Oregon Streams, U.S. Dept Agric. For Ser. General Technical Report 56.
Sullivan, TP., Sullivan, DS. (2001). Influence of variable retention harvest on forest ecosystems. II Diversity and population dynamics of small mammals. Journal of Applied Ecology 38: 1234-1252
Ulbrichová, I., Janeček, V., Štícha, V., Zahradník, D., Kovaříková, M., Topaçoglu, O. (2016). Dead wood structure and volume in managed Fagus orientalis forests in the Black Sea region, Austrian Journal of Forest Science 133: 111-138
Ulikzka, H., Angelstam, P. (2000). Assessing conservation values of forest stands based on specialised lichens and birds. Biological Conservation 95:3 43-351.
Utschick, H. (1991). Beziehungen zwischen Totholzreichtum und Vogelwelt in Wirtschaftswäldern. Forstwissenschaftliches Centralblatt 110 (2): 135-148.
Weggler, K., Dobbertin, M., Jüngling, E., Kaufmann, E., Thürig, E. (2012). Dead wood volume to dead wood carbon: the issue of conversion factors. European Journal of Forest Research 131: 1423-1438.



