Abstract
Few study examined the possible influence of artificially altering soil moisture content on stem respiration (Rs) and stem chemical composition together, we artificially simulated four soil moisture gradients in Dalbergia odorifera (D. odorifera) plantations and investigated Rs, predawn shoot water potential (ψpd), nitrogen content, and non-structural carbohydrates (NSCs) over time. Stem temperature (Ts) and Rs declined continuously during the dry period and reached their minimum values in January. Irrigation increased the Rs rate, while drought decreased it. The nitrogen content of the sapwood remained constant until December, after which it decreased when winter began. During the winter, the sapwood nitrogen increased under irrigation but decreased under drought conditions. The soluble sugars tended to increase during the dry period at the cost of starch, which caused a small but non-significant increase in NSCs. The effect of water treatments on total NSCs was limited; however the drought treatments increased the soluble sugars content but decreased the starch content. Rs explained 49, 72 and 40% of the seasonal variation in the stem nitrogen, starch and soluble sugars, respectively. The ψpd significantly explained 49, 79 and 85% of the variation in the Rs, stem starch and soluble sugars, respectively. Thus, soil moisture content likely affects Rs by affecting the amount of sapwood nitrogen, starch and soluble sugars. Low soil moisture content induced by drought enhanced the transformation of starch to soluble sugars during the winter, when heartwood formed. These strong relationships may provide interesting implication for future study on heartwood formation.
1. Introduction
Dalbergia odorifera T. Chen (D. odorifera) is a medium-sized evergreen tree of the Leguminosae family, which is famous for its heartwood. The heartwood of D. odorifera, named “Jiang Xiang” in traditional Chinese medicine, has been included in the Chinese Pharmacopoeia for centuries. The heartwood of D. odorifera is widely used to stop bleeding, regulate the ‘‘Qi’’, dissipate blood stasis, and relieve pain (Tao & Wang 2010). D. odorifera has been widely used not only as a kind of medicinal material in the pharmaceutical industry but also as a preferred material in luxury furniture and crafts. However, the heartwood of D. odorifera forms relatively slowly; thus, research has focused on the promotion of heartwood formation (Cui et al. 2016).
Stem respiration (Rs) is an important component of tree physiology, accounting for 5 ~ 40% of total annual respiration among different forests (Maseyk et al. 2008, Wieser et al. 2009, Guidolotti et al. 2013). Some field studies have suggested that Rs declines with decreasing soil water content (Levy and Jarvis 1998, Stahl et al. 2011), but not others (Zach et al. 2010, Guidolotti et al. 2013). Discrepancies may arise from the distinct sensitivity of species to water deficit and the lack of studies examining the relationship between tree water availability and Rs. In addition, studies on the effects of irrigation on Rs in the dry season are limited (Maier 2001).
Rs is often correlated with sapwood nitrogen content (Ryan 1991, Vose and Ryan 2002) probably because protein turnover and other processes related to protein metabolism. However, a relationship between Rs and nitrogen content is not always observed in field grown trees. Lavigne and Ryan (1997) found no relationship between Rs and sapwood nitrogen content in several boreal tree species. In contrast, winter stem Rs was correlated with sapwood nitrogen content in eastern white pine (Pinus strobus L.) trees (Vose and Ryan 2001). Experimental data examining effects of soil moisture content on sapwood nitrogen content is limited. Sapwood nitrogen content decreased significantly following drought (Rodríguez-Calcerrada et al. 2014), and it was also observed in stems of beech (Fagus sylvatica L.) seedling (Peuke and Rennenberg 2004).
Non-structural carbohydrates (NSCs) can account for up to 20% stem tissue biomass (Kozlowski 1992). NSCs in stem tissue are intrinsically linked to many physiological processes in plants, and may fluctuate considerably in response to variations in respiration (Pregitzer et al. 2000), however little is known on the relationship between Rs and NSCs in stem tissue (Rodríguez-Calcerrada et al. 2014). Although there are a few cases where tree NSCs show a decline in response to drought (Körner 2003, Anderegg 2012), observations of increased NSCs during drought are much more common in the literature (Würth et al. 2005, Sala and Hoch 2009, Galvez et al. 2011). Experimental studies on seedlings have indicated that an increase in nonstructural carbohydrates (NSCs) in plant bodies enhances their drought tolerance (Secchi and Zwieniecki 2011, O’Brien et al. 2014). However, the contribution of NSCs to drought tolerance in mature trees remains controversial, because studies monitoring variations in NSCs and xylem embolism over time are lacking.
To our knowledge, few study examined the possible influence of artificially altering soil moisture content on all of these variables together. In this study, we measured dynamic changes in Rs, water potential, nitrogen content, and NSCs in contrasting soil moisture conditions continuously during the dry season from 2016 to 2017. The objectives of this study were to (1) investigate the variation patterns of Rs, water potential, nitrogen content, and NSCs (2) present new data on the effects of altered soil moisture content on Rs, water potential, nitrogen content, and NSCs in stem tissue during the dry period; (3) determine the possible relationships among Rs, water potential, nitrogen content, and NSCs.
2. Materials and methods
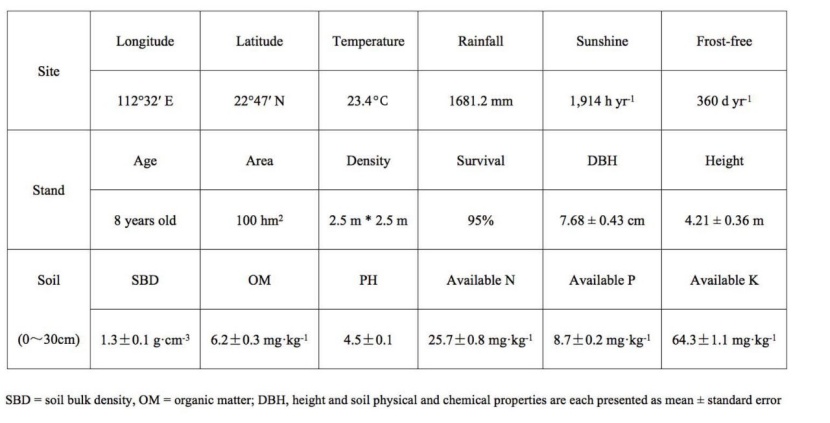
Table 1: Climate, forest conditions and soil physical and chemical properties of the study region
2.1 Plant material and study area
The study site was a young 8-year-old plantation of D. odorifera, when the heartwood formation begins. The plantation was located in Foshan, Guangdong Province, in southern China (22°47´N, 112°32´E). The mean annual precipitation is 1681.2 mm, with 562 mm occurring in the dry season (from October to March). The mean annual air temperature is 23.4 °C, with the minimum in January (19.3 °C) and the maximum in July (33.8 °C). In 2009, 1-year-old seedlings of D. odorifera were bought from Hainan Island (the origin) and the plantation was established using an approximately 2.5-m × 2.5-m planting grid. The plantation presented a survival rate of 95% and had a 7.68 ± 0.43 cm mean stem diameter at breast height (DBH) at the beginning of treatments and 4.21 ± 0.36 m mean tree height. For Rs measurement, two trees were selected in the central region of each plot, i.e., six sampled trees of each regime were chosen in the middle of three plots. The mean DBH at the beginning of treatments was 8.8 ± 0.7 cm for HD trees, 8.5 ± 0.4 cm for LD trees, 9.0 ± 0.8 cm for DI trees, and 8.4 ± 0.5 cm for CK trees. The slope of plots was approximately 15°, and ground vegetation was dominated by Polygonum chinense and Bidens pilosa. The soil is a latosol (developed from granite) with a depth > 1 m. Characteristics of the site are listed in Table 1.
2.2 Experimental design and soil moisture content
The experiment was a completely randomized block design. To widen the range of the soil moisture status during the dry season, we arranged four regimes with three blocks, resulting in a total of 12 plots of 400 m2 each: (1) heavy drought (HD), in which ≈74.8 % of rainfall was intercepted by 2-m-diameter polyvinyl chloride (PVC) rain gutters that conducted water away from three plots; (2) light drought (LD), in which ≈46.1 % of rainfall was intercepted by 1.3-m-diameter PVC rain gutters that conducted water away from three plots; (3) dry season irrigation (DI), in which an amount of water equal to the monthly rainfall water was intercepted by the HD setup and diverted to three plots for irrigation from October 2016 on; (4) and the control (CK). The four treatment plots of the same block were 5 meters apart from each other. The throughfall exclusion system has been described previously (Limousin et al. 2009, Rodríguez-Calcerrada et al. 2011). In each plot, there is about 60 trees, and eight rain gutters of 20 meters long were installed one meter high from the ground. To avoid the lateral water movement into the dried out plot as possible, all plots were chosen on the upper slopes and each plot was separated by water-stop sheets of 3 meters long and 2 meters wide. Soil volumetric moisture content was monitored by a soil moisture monitoring system (AZS-2C, TDR, China) at a depth of ≈ 30 cm once per hour. In each plot, three sensors were installed in 3 different positions. Each sensor was installed 1 m far from the tree base in the same direction under the gutters.
2.3 Stem respiration
The Rs at 1.3 m was measured in situ using an LI-8100 CO2 flux system (Li-Cor Inc., Lincoln, NE, USA) in 2-week intervals as described previously by Xu et al. (2000). A custom-built cylindrical polyvinyl chloride (PVC) collar of 8 cm in diameter and 10 cm in height was attached to the stem surface to connect the stem and the LI-8100 CO2 flux system. The two ends of the opaque PVC collar were cut to match the approximate curvature of the stem and the LI-8100 CO2 flux system. The collar was attached to the stem using silicone sealant, and was maintained in the same location throughout the study period. The measurements (randomization between treatments, block by block) occurred from 9:00 to 12:00 from October 2016 to March 2017(The sunrise is between 6:40 and 7:10). Ts at a depth of 1 cm was also measured simultaneously at each sampling location using a portable electronic thermometer, the thermometer has a sharp probe, which can make sure good contact with the wood. The Rs and Ts were uniformly measured on south-facing stems in this study.
The Q10 values were used as a parameter of temperature sensitivity of the stem with the following the equation:
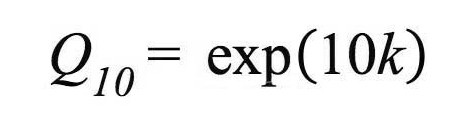
with "k " being the slope of the regression equation of Rs against Ts (Atkin et al. 2005).
2.4 Water potential
On the same days of every Rs measurement in 2-week intervals, one shoot from each tree in which the Rs was measured was excised before dawn and immediately measured using a pressure chamber (PMS Instrument Company, Albany, OR, USA).
2.5 Chemical composition
To avoid affecting the measurement of Rs, stem cores (the outer 1 cm of sapwood) at breast height of 4 ~ 5 surrounding trees belonging to each treatment were collected by a stem corer every two weeks. Samples were collected between 9:00 and 12:00 to minimize the diel variation in NSCs, we immediately chilled them on ice, and then microwaved them for 90 s at 600 W within 2 h of collection to stop carbohydrate enzymatic reactions (Hoch et al. 2003), after which the samples were oven-dried at 65 °C to a constant weight. We ground the dried tissues to a fine powder and stored at 4 °C until analyses. The nitrogen content was measured using an elemental analyzer (Flash EA1112, USA). The NSCs (soluble sugars and starch) were analyzed using the enzymatic assay of McCleary et al. (1997) as described previously by Yang et al. (2015).
2.6 Statistical analysis
The mean values and standard errors were calculated and analyzed. The equations for the calculations and the plots for the graphs were generated in Microsoft Excel 2010 (USA); the analysis of variance and regression analysis were performed using the data processing software SPSS 17.0 (IBM, NY, USA).
3. Results
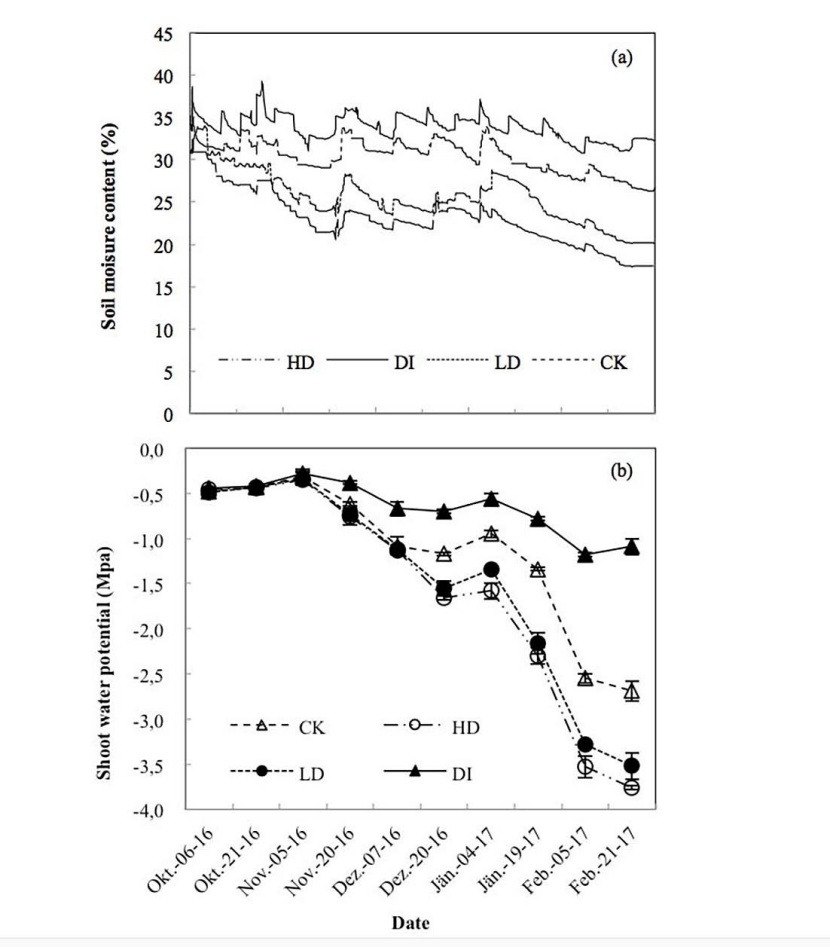
Figure 1: Soil moisture content (a) and predawn shoot water potential (ψpd) (b) during the dry period (2016-2017). Values of ψpd are the means ± SE (n = 6).
3.1 Soil moisture content and shoot water potential
An overall decreasing trend in soil moisture content was observed during the dry period, with the minimum values occurring in February (Figure 1a). The soil moisture content was significantly lower for the HD and LD treatments subjected to throughfall exclusion but higher for DI (P < 0.01). This decrease in soil moisture content caused continuous decline in the shoot water potential (ψpd) during the dry period. Significant differences in soil moisture content caused significant differences (P < 0.01) in the shoot water potential (ψpd) among water treatments from the end of November to February (Figure 1b).
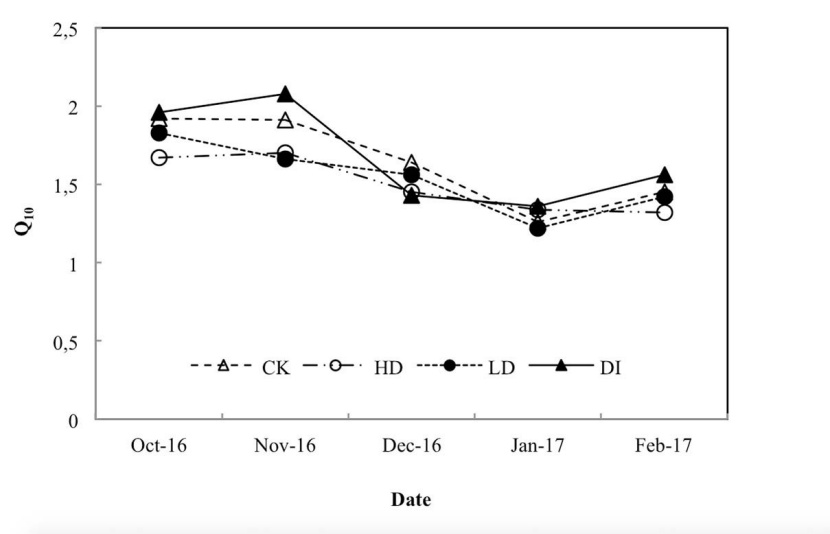
Figure 2: Seasonal variation of sensitivity of stem respiration (Rs) to variations in temperature (Q10).
3.2 Stem temperature and respiration
The results showed that the values of Q10 varied over the dry season without a clear pattern (Figure 2). Ts decreased continuously during the dry period and reached its minimum value in January (Figure 3a). We found no significant differences in Ts among water treatments except in December and January (P < 0.05), which are in the coldest period of the year (Figure 3a). The seasonal variation patterns of Rs were similar to those of Ts (Figure 3b). From the end of October, Rs significantly differed among water treatments (Figure 3b). We found that the Rs differed significantly (P < 0.05) among water treatments, following the order DI > CK > LD > HD (Figures 3b).
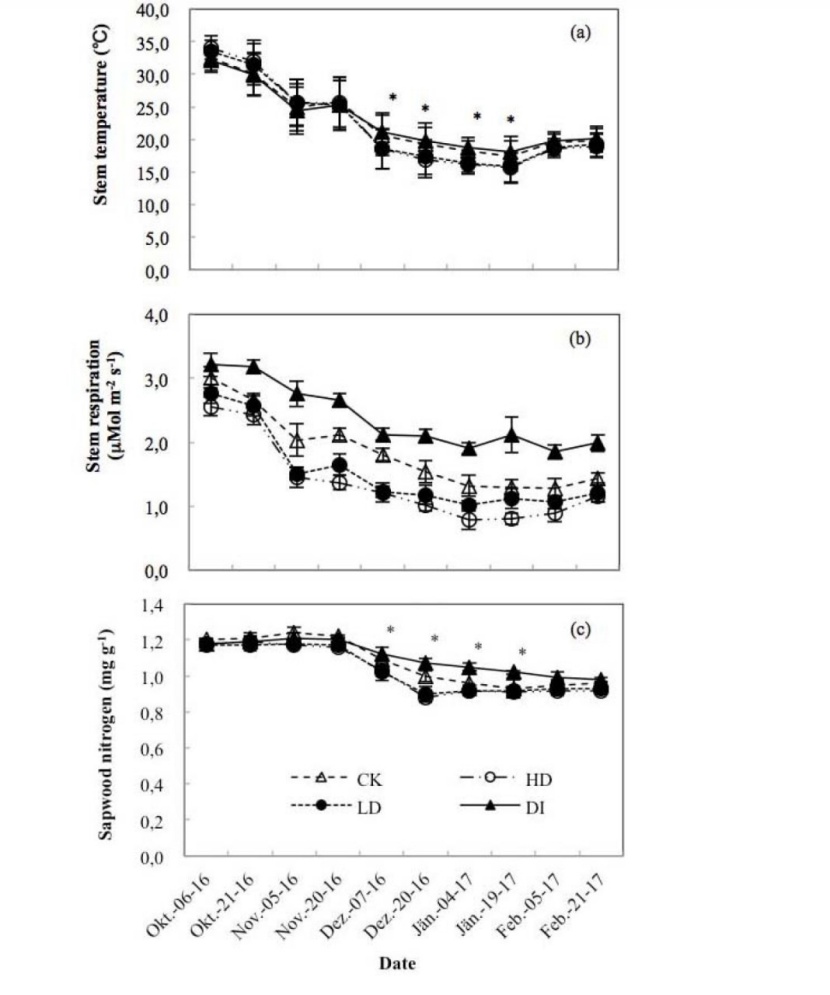
Figure 3: Stem temperature (Ts) (a), respiration (Rs) (b) and nitrogen content (c) during the dry period. Values are the means ± SE (n = 3-6). “*” indicates significant differences (P < 0.05) among the CK, HD, LD and DI treatments
3.3 Chemical composition
The sapwood nitrogen content remained constant until December, after which it decreased as winter began. Significant differences (P < 0.05) were found in sapwood nitrogen content among water treatments in December and January (Figure 3c). A decreasing trend for sapwood starch (Figure 4a) and an increasing trend for soluble sugars (Figure 4b) were observed through the sampling dates, which caused a small but non-significant increase (P > 0.05) in the total NSCs (Figure 4c). Significant variations in starch and soluble sugars were observed among water treatments; however, there were no significant differences between DI and CK or HD and LD (Figure 4a, 4b). No significant difference was observed in the total NSCs among treatments (Figure 4c).
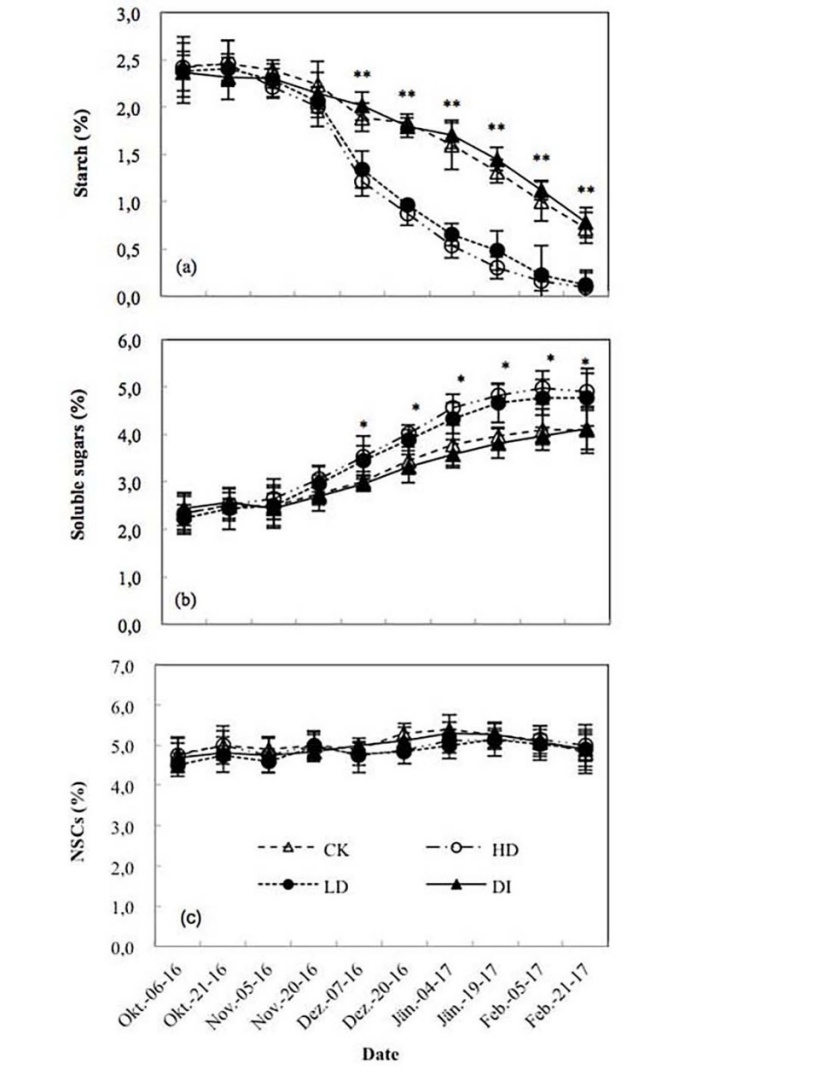
Figure 4: The concentrations of starch (a), soluble sugars (b) and non-structural carbohydrates (NSCs) (c) in the sapwood during the dry period. Values are the means ± SE (n = 3). “%” means % dry mass of sapwood. “*” and “**” indicate significant differences at the 0.05 and 0.01 levels, respectively, among the CK, HD, LD and DI treatments.
3.4 Relationship among water potential, respiration and chemical composition
The seasonal pattern of Rs was related to the seasonal changes in stem nitrogen, starch and soluble sugars. Rs explained 49, 72 and 40% of the seasonal variation in the stem nitrogen, starch and soluble sugars, respectively (Figure 5). The variation in ψpd significantly (P < 0.05) explained 49, 79 and 85% of the variation in Rs, starch and soluble sugars during the dry period, respectively (Figure 6).

Figure 5: Relationships between stem respiration (Rs) and the concentrations of nitrogen (a), starch (b) and soluble sugars (c) in the sapwood. “%” means % dry mass of sapwood. P < 0.05 for all regression equations.
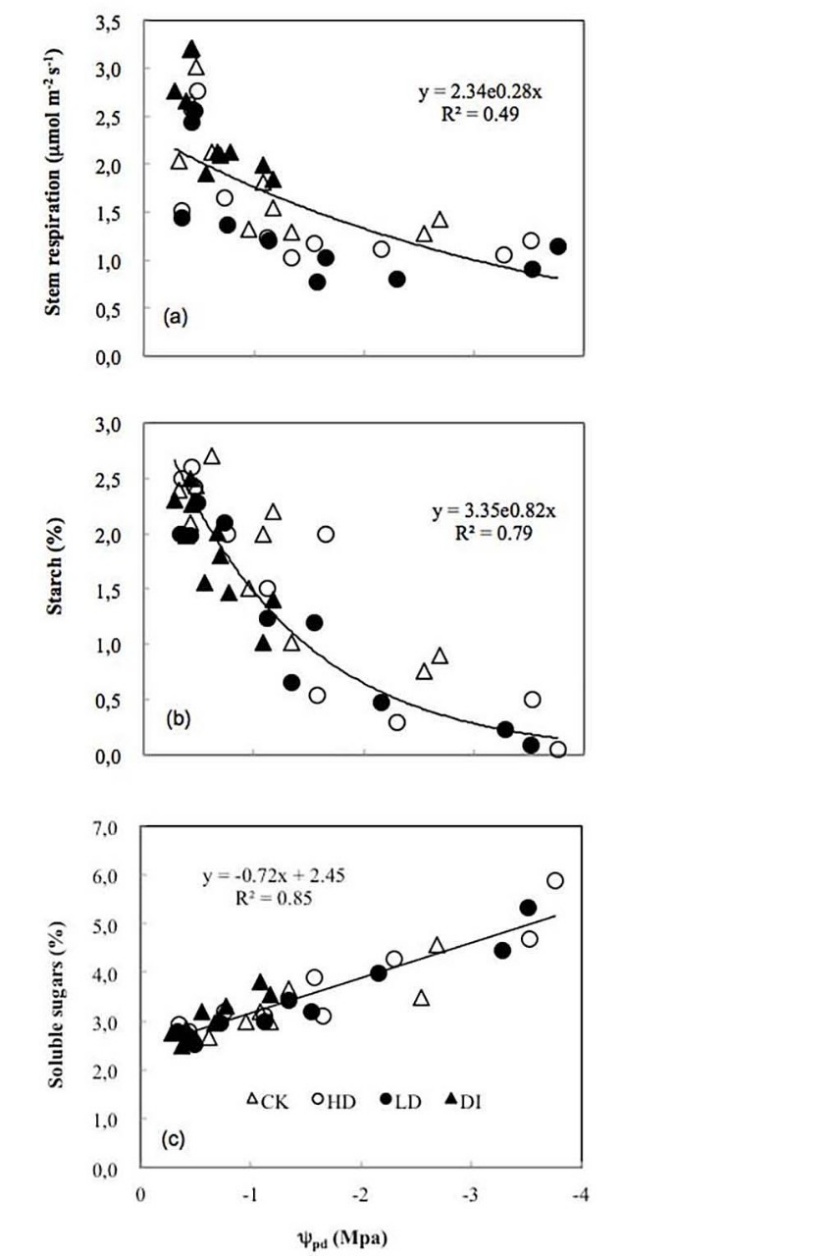
Figure 6: Relationships among predawn shoot water potential (ψpd) and stem respiration (a), starch (b) and soluble sugars (c). “%” means % dry mass of sapwood. P < 0.05 for all regression equations.
4. Discussion
4.1 Seasonal changes in stem respiration and chemical composition
The similar seasonal pattern of moisture condition of soil and trees indicate that soil moisture content had a significant and positive effect on the moisture condition of trees (Granda et al. 2014, Rodríguez-Calcerrada et al. 2014). During the dry season, the decreasing temperature decreased stem respiration by inhibiting the enzyme activity (Gaumont-Guay et al. 2006). The sapwood nitrogen content decreased as winter began. Maier (2001) also observed sapwood nitrogen to decrease with Ts during the winter. This decrease in sapwood nitrogen was likely caused by a progressive reduction in the demand for energy from physiological processes. A decreasing trend for sapwood starch and an increasing trend for soluble sugars were consistent with the data of Terziev et al. (1997), which caused a small but non-significant increase (P > 0.05) in the total NSCs (Hoch et al. 2003). Soluble sugars in the sapwood tended to increase during the dry period at the cost of starch (Sevanto et al. 2014).
4.2 Changes in stem respiration and chemical composition in response to altered soil moisture content
Hartmann et al. (2013a) observed that drought could reduce whole-plant respiration. Drought can also slow Rs rates by reducing xylem CO2 concentrations (Teskey and McGuire 2005; Saveyn et al. 2007a), respiratory substrate availability (Wertin and Teskey 2008; Maier 2001), phloem transport (Sevanto et al. 2014), and energy demands of cell maintenance processes (Atkin & Macherel 2009). Drought-induced low turgor pressure of living cells (Zweifel et al. 2006) could also be a factor underpinning low Rs (Saveyn et al. 2007a, 2007b). However, studies on the effects of irrigation on Rs are limited. Maier et al. (1998) observed that Rs increased in response to irrigation and fertilization treatments during the dormant season; however, irrigation did not have a significant effect on Rs during the growth season (Maier 2001). These results were consistent with our observation that DI increased the Rs rate during the dry period. During the same period, low soil moisture content was the main factor limiting the physiology of the trees, but this factor was eliminated in the DI treatment. In addition, a number of mechanisms maintaining stem functionality might regulate respiratory metabolism (Rodríguez-Calcerrada 2014). Thus, the irrigation-induced increase in Rs was likely caused by a progressive increase in the energy demand from physiological processes.
Interestingly, we found no significant differences in either Ts or sapwood nitrogen content among water treatments except in December and January (P < 0.05), which are in the coldest period of the year (Figure 3a, 3c). HD and LD trees with lower water potential were more vulnerable to sudden drops in temperature in December and January, and this vulnerability resulted in significantly lower Ts values for trees in the HD and LD treatments compared with those for trees in the CK and DI treatments. A positive correlation exists between Ts and sapwood nitrogen in winter (Maier 2001). Ultimately, significant differences in Ts caused by sudden temperature drops resulted in significant differences in sapwood nitrogen during the winter.
In this study, the effects of water treatments on the total NSCs were very limited; however drought treatments increased soluble sugars but decreased the starch content. Observations of NSC concentration changes in trees during drought are rather inconclusive, though; concentrations may increase or decrease or not change at all (Galvez et al. 2011, Poyatos et al. 2013, Mitchell et al. 2013, Rodríguez-Calcerrada et al. 2014) and this response can even vary across plant organs of individual trees (Hartmann et al. 2013b). The phenomenon that drought increased soluble sugars but decreased the starch content has also been observed by Sala and Hoch (2009) and Rodríguez-Calcerrada et al. (2014).
4.3 Relationship between stem respiration and chemical composition
The decrease in stem nitrogen coincided with reductions in Rs during the dry period. The relationship between Rs and stem nitrogen varies for other species. Rs is not related to stem nitrogen in Picea abies (Stockfors & Linder 1998) or in several other boreal species (Lavigne & Ryan 1997). However, Ryan (1991) showed that respiration in several plant tissues increased linearly with tissue nitrogen. Rs also increased with stem nitrogen in Pinus strobus (Vose & Ryan 2002) and loblolly pine (Maier 2001). If the stem nitrogen is indicative of protein content (Penning de Vries 1975) and protein metabolism consumes the largest portion of Rs in woody tissues (Maier 2001), then changes in nitrogen might result in significant changes in Rs. In our study, Rs varied with stem nutrition. This variance may cause serious errors when estimating the Rs of D. odorifera stands growing under different nutrient regimes; thus, other methods of estimating stand Rs may be needed.
NSCs are intrinsically linked to many physiological processes in plants and can fluctuate considerably in response to variations in Rs (Pregitzer et al. 2000). Rs is significantly related to the soluble sugar content in the leaves and roots of many tree species (Atkin and Tjoelker 2003, Kruse et al. 2011) but has rarely been examined in stems (Gansert et al. 2002). To our knowledge, few studies have compared the effects of drought or irrigation on Rs together with NSCs (see Rodríguez-Calcerrada et al. 2014). Rodríguez-Calcerrada et al. (2014) did not find a significant relationship between Rs and NSCs. In our study, Rs explained 72 and 40% of the variation in the starch content and soluble sugar content during the dry period, respectively. This discrepancy might be attributed to differences in the study period and tree species. Future studies comparing the effects of drought or irrigation on Rs together with NSCs are warranted. Based on the strong relationships among Rs, sapwood nitrogen, starch and soluble sugars, soil moisture content likely affects Rs by affecting the sapwood nitrogen, starch and soluble sugars.
4.4 Interesting implication for future study
As is well-known, heartwood formation is a secondary metabolic process. “Jiang Xiang”, the heartwood of D. odorifera, contains volatile oils and flavonoids. These secondary metabolites are likely reflected by NSC accumulation because of decreased growth and sink activity (Herms & Mattson 1992). In other words, NSCs bridge the conversion of photosynthetic products to secondary metabolites through the transformation of starch to soluble sugars (Hartmann & Trumbore 2016). Abundant evidence has suggested that heartwood forms during the dormant or dry season (Shain & Hillis 1973, Nelson 1978, Beauchamp et al. 2013), when stems require more secondary metabolic substrates obtained by the transformation of starch to soluble sugars. Meng et al. (2010) investigated D. odorifera in multiple locations and observed that drought could promote heartwood formation in D. odorifera. In our study, low soil moisture content induced by drought enhanced the transformation of starch to soluble sugars during winter. These strong relationships could provide data to support the hypothesis that drought promotes heartwood formation in D. odorifera. Further studies on the effects of drought on altered NSC levels in different zones of the stems and the possible relationships of these phenomena with heartwood formation are warranted. In addition, based on the significant relationship between Rs and NSCs (starch and soluble sugars), variation in the Rs rate may also indicate changes in heartwood formation during the dry period.
5. Conclusions
In addition to total NSCs, significant seasonal variation could be observed in Rs, water potential, nitrogen content, starch and soluble sugars. Irrigation increased while drought decreased Rs. Drought only affected nitrogen content during the winter. The effect of soil moisture content on total NSCs was limited; however drought enhanced the transformation of starch to soluble sugars during the dry period. These strong relationships among Rs, water potential, nitrogen content, starch and soluble sugars may provide interesting implication for future study on heartwood formation. Further studies on the effects of drought on the radial distribution of NSC levels in the stems and the possible relationships of these phenomena with heartwood formation are warranted.
Acknowledgements
This study was supported by the research funds (2016YFD060020503, CAFYBB2016ZX003-4) and the National Natural Science Foundation of China (grant nos. 31470026 and 30872052). Special thanks are extended to the editors and the anonymous reviewers for their constructive suggestions and comments.
References
ANDEREGG WR. 2012. Complex aspen forest carbon and root dynamics during drought. Climatic Change 111: 983-991.
ATKIN OK, BRUHN D & TJOELKER MG. 2005. Response of plant respiration to changes in temperature: mechanisms and consequences of variations in Q10 values and acclimation. In: Lambers H, Ribas-Carbó M. (Eds.), Plant Respiration. From Cell to Ecosystem. Springer, Dordrecht, The Netherlands, pp. 95-135.
ATKIN OK & MACHEREL D. 2009. The crucial role of plant mitochondria in orchestrating drought tolerance. Annals of Botany 103: 581-597.
ATKIN OK & TJOELKER MG. 2003. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends in Plant Science 8: 343-351.
BEAUCHAMP K, MENCUCCINI M, PERKS M & GARDINER B. 2013. The regulation of sapwood area, water transport and heartwood formation in Sitka spruce. Plant Ecology & Diversity 6: 45-56.
CUI ZY, XU DP, YANG ZJ, ZHANG NN, LIU XJ & HONG Z. 2016. A review of mechanism and artificial promotion of heartwood formation. World Forestry Research 29: 33-37.
GALVEZ DA, LANDHÄUSSER SM & TYREE MT. 2011. Root carbon reserve dynamics in aspen seedlings: does simulated drought induce reserve limitation? Tree Physiology 31: 250-257.
GANSERT D, BACKES K, OZAKI T & KAKUBARI Y. 2002. Seasonal variation of branch respiration of a treeline forming (Betula ermanii Cham.) and a montane (Fagus crenata Blume) deciduous broad-leaved tree species on Mt. Fuji, Japan. Flora 197: 186-202.
GAUMONT-GUAY D, BLACK TA, GRIFFIS TJ, BARR AG, MORGENSTERN K, JASSAL RS & NESIC Z. 2006. Influence of temperature and drought on seasonal and interannual variations of soil, bole and ecosystem respiration in a boreal aspen stand. Agriculture and Forest Meteorology 140: 203-219.
GRANDA V, DELATORRE C, CUESTA C, CENTENO ML, FERNÁNDEZ B, RODRÍGUEZ A & FEITO I. 2014. Physiological and biochemical responses to severe drought stress of nine Eucalyptus globulus clones: a multivariate approach. Tree Physiology 34: 778-786.
GUIDOLOTTI G, REY A, D’ANDREA E, MATTEUCCI G & DE ANGELIS P. 2013. Effect of environmental variables and stand structure on ecosystem respiration components in a Mediterranean beech forest. Tree Physiology 33: 960-972.
HARTMANN H & TRUMBORE S. 2016. Understanding the roles of nonstructural carbohydrates in forest trees-from what we can measure to what we want to know. New Phytologist 211: 386-403.
HARTMANN H, ZIEGLER W, KOLLE O & TRUMBORE S. 2013a. Thirst beats hunger declining hydration during drought prevents carbon starvation in Norway spruce saplings. New Phytologist 200: 340-349.
HARTMANN H, ZIEGLER W & TRUMBORE S. 2013b. Lethal drought leads to reduction in nonstructural carbohydrates in Norway spruce tree roots but not in the canopy. Functional Ecology 27: 413–427.
HERMS DA & MATTSON WJ. 1992. The dilemma of plants: to grow or defend. Quarterly Review of Biology 67: 283-335.
HOCH G, RICHTER A & KORNER C. 2003. Non-structural carbon compounds in temperate forest trees. Plant Cell and Environment 26: 1067-1081.
KÖRNER C. 2003. Carbon limitation in trees. Journal of ecology 91: 4-17.
KOZLOWSKI TT. 1992. Carbohydrate sources and sinks in woody plants. The Botanical Review 58: 107-222.
KRUSE J, RENNENBERG H & ADAMS MA. 2011. Steps towards a mechanistic understanding of respiratory temperature responses. New Phytologist 189: 659-677.
LAVIGNE MB & RYAN MG. 1997. Growth and maintenance respiration rates of aspen, black spruce and jack pine stems at northern and southern BOREAS sites. Tree Physiology 17: 543-551.
LEVY PE & JARVIS PG. 1998. Stem CO2 fluxes in two Sahelian shrub species (Guiera senegalensis and Combretum micranthum). Functional Ecology 12: 107-116.
LIMOUSIN JM, RAMBAL S, OURCIVAL JM, ROCHETEAU A, JOFFRE R & RODRÍGUEZCORTINA R. 2009. Long-term transpiration change with rainfall decline in a Mediterranean Quercus ilex forest. Global Change Biology 15: 2163-2175.
MAIER CA. 2001. Stem growth and respiration in loblolly pine plantations differing in soil resource availability. Tree Physiology 21: 1183-1193.
MAIER CA, ZARNOCH SJ & DOUGHERTY PM. 1998. Effects of temperature and tissue nitrogen on dormant season stem and branch maintenance respiration in a young loblolly pine (Pinus taeda) plantation. Tree Physiology 18: 11-20.
MASEYK K, GRÜNZWEIG JM, ROTENBERG E & YAKIR D. 2008. Respiration acclimation contributes to high carbon use efficiency in a seasonally dry pine forest. Global Change Biology 14: 1553-1567.
MCCLEARY BV, GIBSON TS & MUGFORD DC. 1997. Measurement of total starch in cereal products by amyloglucosidase–alpha-amylase method: collaborative study. Journal of Aoac International 80: 571-579.
MENG H, XIE C, YANG Y, WEI J & FENG J. 2010. Suitable producing areas of Dalbergia odorifera T. Chen. Lishizhen Medicine & Materia Medica Research 9: 2304-2306.
MITCHELL PJ, O’GRADY AP, TISSUE DT, WHITE DA, OTTENSCHLAEGER ML & PINKARD EA. 2013. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytologist 197: 862-872.
NELSON ND. 1978. Xylem ethylene, phenol-oxidizing enzymes, and nitrogen and heartwood formation in walnut and cherry. Canadian Journal of Botany 56: 626-634.
O’BRIEN MJ, LEUZINGER S, PHILIPSON CD, TAY J & HECTOR A. 2014. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nature Climate Change 4: 710-714.
PENNING DE & VRIES FWT. 1975. The cost of maintenance processes in plant cells. Annals of Botany 39: 77-92.
PEUKE AD & RENNENBERG H. 2004. Carbon, nitrogen, phosphorus, and sulphur concentration and partitioning in beech ecotypes (Fagus sylvatica L.): phosphorus most affected by drought. Trees 6: 639-648.
POYATOS R, AGUADE D, GALIANO L, MENCUCCINI M & MARTINEZ-VILALTA J. 2013. Drought-induced defoliation and long periods of near-zero gas exchange play a key role in accentuating metabolic decline of Scots pine. New Phytologist 200: 388–401.
PREGITZER KS, KING JS, BURTON AJ & BROWN SE. 2000. Response of tree fine roots to temperature. New Phytologist 147: 105-115.
RODRÍGUEZ-CALCERRADA J, JAEGER C, LIMOUSIN JM, OURCIVAL JM, JOFFRE R & RAMBAL S. 2011. Leaf CO2 efflux is attenuated by acclimation of respiration to heat and drought in a Mediterranean tree. Functional Ecology 25: 983-995.
RODRÍGUEZ-CALCERRADA J, MARTIN-STPAUL NK, LEMPEREUR M, OURCIVAL JM, DEL REY M DC, JOFFRE R & RAMBAL S. 2014. Stem CO2 efflux and its contribution to ecosystem CO2 efflux decrease with drought in a Mediterranean forest stand. Agricultural and Forest Meteorology 195: 61-72.
RYAN MG. 1991. Effects of climate change on plant respiration. Ecological Applications 1: 157-167.
SALA A & HOCH G. 2009. Height-related growth declines in ponderosa pine are not due to carbon limitation. Plant Cell and Environment 32: 22-30.
SAVEYN A, STEPPE K & LEMEUR R. 2007a. Drought and the diurnal patterns of stem CO2 efflux and xylem CO2 concentration in young oak (Quercus robur). Tree Physiology 27: 365-374.
SAVEYN A, STEPPE K & LEMEUR R. 2007b. Daytime depression in tree stem CO2 efflux rates: is it caused by low stem turgor pressure? Annals of Botany 99: 477-485.
SECCHI F & ZWIENIECKI MA. 2011. Sensing embolism in xylem vessels: the role of sucrose as a trigger for refilling. Plant, Cell & Environment 34: 514-524.
SEVANTO S, MCDOWELL NG, DICKMAN T, PANGLE R & POCKMAN WT. 2014. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell and Environment 37: 153-161.
SHAIN L & HILLIS WE. 1973. Ethylene production in xylem of Pinus radiata in relation to heartwood formation. Canadian Journal of Botany 51: 1331-1335.
STAHL C, BURBAN B, GORET JY & BONAL D. 2011. Seasonal variations in stem CO2 efflux in the Neotropical rainforest of French Guiana. Annals of Forest Science 68: 771-782.
STOCKFORS J & LINDER S. 1998. Effect of nitrogen on the seasonal course of growth and maintenance respiration in stems of Norway spruce trees. Tree Physiology 18: 155-166.
TAO Y & WANG Y. 2010. Bioactive sesquiterpenes isolated from the essential oil of Dalbergia odorifera T. Chen. Fitoterapia 81: 393-396.
TERZIEV N, BOUTELJE J & LARSSON K. 1997. Seasonal fluctuations of low-molecular-weight sugars, starch and nitrogen in sapwood of Pinus sylvestris L. Scandinavian Journal of Forest Research 12: 216-224.
TESKEY RO & MCGUIRE MA. 2005. CO2 transported in xylem sap affects CO2 efflux from Liquidambar styraciflua and Platanus occidentalis stems, and contributes to observed wound respiration phenomena. Trees-Structure and Function 19: 357-362.
VOSE JM & RYAN MG. 2002. Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis. Global Change Biology 8: 182-193.
WERTIN TM & TESKEY RO. 2008. Close coupling of whole-plant respiration to net photosynthesis and carbohydrates. Tree Physiology 21: 1831-1840.
WIESER G, GRUBER A, BAHN M, CATALÁ E, CARRILLO E, JIMÉNEZ MS & MORALES D. 2009. Respiratory fluxes in a Canary Islands pine forest. Tree Physiology 29: 457-466.
WÜRTH MK, PELAEZ-RIEDL S, WRIGHT SJ & KÖRNER C. 2005. Non-structural carbohydrate pools in a tropical forest. Oecologia 143: 11-24.
XU M, DEBIASE TA & QI Y. 2000. A simple technique to measure stem respiration using a horizontally oriented soil chamber. Canadian Journal of Forest Research 30: 1555-1560.
YANG Q, LIU L, ZHANG W, XU M & WANG S. 2015. Different responses of stem and soil CO2 efflux to pruning in a Chinese fir (Cunninghamia lanceolata) plantation. Trees-Structure and Function 29: 1207-1218.
ZWEIFEL R, ZIMMERMANN L, ZEUGIN F & NEWBERY DM. 2006. Intra-annual radial growth and water relations of trees: implications towards a growth mechanism. Journal of Experimental Botany 6: 1445-1459.



