Abstract
Disturbance regimes and gap characteristics of the forest stands are the conceptual foundations for the core of silviculture. The gap characteristics and disturbance regime were investigated in an intact old-growth oriental beech stand in a forest reserve of Hyrcanian region, north of Iran. Seventy six canopy gaps were sampled at 5 transects 625 to 2000 m. The gap area was estimated using two field methods of measurement: elliptical (ELLIP) and non-convex polygonal (NOPO). The mean canopy gap fraction was 8.4% and 10.8% based on NOPO and ELLIP, respectively. The mean expanded gap fraction calculated by ELLIP (22.0%) was greater than that by NOPO (18.1%). Mean canopy gap size was 159 m2 and 208.7 m2 for NOPO and ELLIP methods, respectively. Mean expanded gap size was 515.7 m2 and 626 m2 for NOPO and ELLIP methods, respectively. Most of canopy gaps (68.4%) were smaller than 200 m2, whereas only 1.4% was bigger than 500 m2. Nearly 90% of number and area of whole expanded gaps belonged to classes of 200-1000 m2. As canopy gap size increased the gap area-perimeter relationship was obviously differed from circular shape, indicating that gap geometry became more complex with gap size. The expanded gaps also followed this pattern, but in expanded gaps (< 450 m2) complexity was less than that in canopy gaps. The maximum of gap-makers was 6 trees per gap but 53% of gaps were formed by the death of two trees or more. Near 43% of gaps were expanded after initial gap formation, due to recurrence of disturbance events. The species composition of gap-makers was similar to those of gap border and forest canopy. We could not identify the mode of mortality of 12.3% gap-makers; however, 60.4% and 19.3% of them were snapped-alive and uprooted, respectively. Exogenous agents such as chronic winds could have a prominent role in initial formation and expansion of gaps.
1. Introduction
Disturbance regimes and stand development processes are the conceptual foundations for the core of silviculture (Kohm and Franklin, 1997). Successful prediction of the effect of a silvicultural manipulation must be based on an understanding of the spatial and temporal dynamics of forest response to different kinds, sizes, frequencies and intensities of disturbances (Smith, 1993). Forest disturbance is a permanent feature of forest ecosystems, determining species composition, structure and process (McCarthy, 2001; Kucbel et al., 2010) and defined as any relatively discrete event in time that disrupts ecosystem, community, or population structure and changes resource pools, substrate availability, or the physical environment (White and Pickett, 1985). Natural disturbances (occurred by winds, insects, diseases, acidic deposition, drought and climate change) with low intensity and relatively high frequency are caused the formation of the gap with different sizes that driving the forest dynamics in the absence of human interventions (Schliemann and Bockheim, 2011).
Forest gap is a space in the canopy produced by the death of at least one over-storey tree where active recruitment of new individuals into the over-storey was occurring (Watt, 1947) and Forest canopy gaps are measurable indicators of past small-scale disturbances (Lundquist and Beatty, 2002). Among several metrics used in describing such disturbance patterns; gap frequency, size, and shape are the most important (Gagnon et al., 2004; Schliemann and Bockheim 2011). Gap size reflects the magnitude of the disturbance, which has a direct influence on gap microclimate, understorey damage levels, nutrient cycling and growth of the gap-border trees as well as vegetation elements within gaps (Gray et al., 2002; Muscolo et al., 2007; De Lima et al., 2008).
Gap shape varies highly and that no single geometric form is able to predict the shape of all gaps; the ellipse, in particular, should no longer be used as a standard gap shape (De Lima et al., 2013). While, nonconvex polygons are used to the geometry of gaps, mapped gap areas are 22% less than estimates based on an elliptical model of gap shape and 22% more than area estimates based on a rhomboidal model of gap shape (Battles et al., 1996). Meanwhile, in the study of forest gaps, the geometry of gaps is the center of attention of researchers due to impact on photosynthetically active radiation (PAR) at the forest floor of gaps (Canham, 1988).The mortality agents responsible for gap formation (endogenous or exogenous) can affect gap fraction and gap size as well as the spatial and temporal pattern of gap formation (Woods, 2004). Large canopy openings can form suddenly during a more intense disturbance or gradually from the expansion and coalescence of smaller gaps (Whitmore, 1989). The repeated disturbance processes, along with delimitation methods, strongly influences the estimation of turnover rate and therefore the interpretation of forest dynamics. These issues demonstrate the importance of further studies on repeated disturbances, which is often neglected in forest studies (De Lima et al., 2008).
Hyrcanian forests with an area 1.9 million ha has been located mainly in the south coasts of the Caspian Sea in northern of Iran by altitudinal gradient of -20 to 2200 meters a.s.l. and ~20000 in the Republic of Azerbaijan. This region is covered with temperate deciduous forests. Oriental beech (Fagus orientalis Lipsky) is the most important commercial species in these forests, which forms pure and mixed stands in a belt between 700 and 2000 m.a.s.l. (Sagheb-Talebi and Schütz, 2002). Historically, implementation of forest management plans in this region is referred to five decades ago. Until to 1995, beech stands have been managed chiefly using the shelterwood system. In the most stands, this method has been replaced by selection system due to the technical reasons and particularly regeneration problems. Managed plans had been prepared for only 60% of the surface of these forests, while undisturbed, unmanaged and natural semi-virgin beech forests are still found within the region (Sagheb-Talebi et al., 2005).
It may be stated that the intact oriental beech stands of the Hyrcanean forests can provide a unique opportunity to study the disturbance regimes of forest ecosystems in the absence of human interventions. For this purpose, in order to gain a better understanding of natural gap disturbance processes and investigating the effect of geometric form of gaps (elliptical or nonconvex polygons) on the gap fraction and gap characteristics of these forests we selected an old-growth oriental beech (Fagus orientalis Lipsky) stand and addressed the following questions:
1) What are the characteristics of the gap disturbance regime (i.e., gap fraction, gap size distribution, and gap shape) which reveal disturbance processes? 2) Does the geometric form of the gap effect on the gap size and gap fraction? 3) Is the mortality process responsible for gap formation associated with endogenous or exogenous disturbance? 4) Are larger gaps formed from the expansion of smaller gaps or during single mortality events?
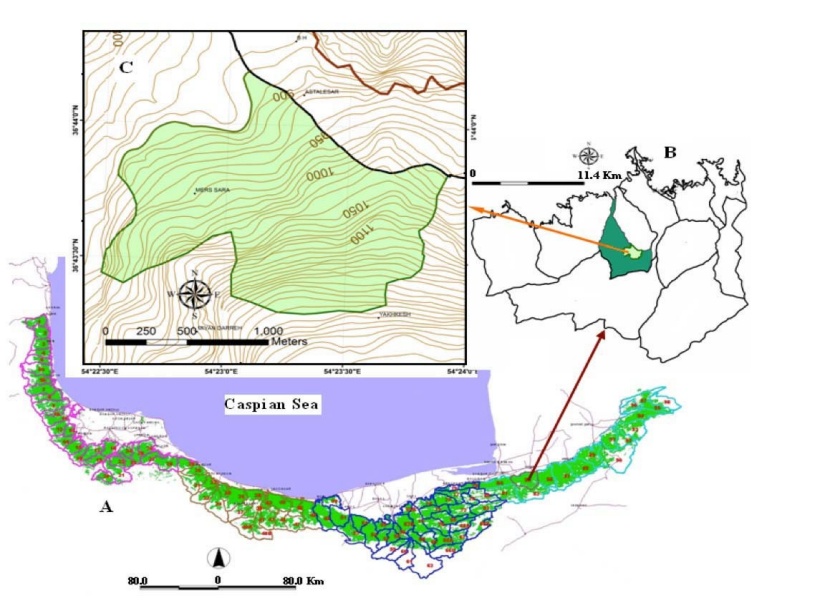
Fig. 1: A) Location watershed 85 in north forests of Iran, B) Location district 2 Shast-Kalate in watershed 85 and C) Location of the study area in district 2 Shast-Kalate
2. 1 Study area
The study was conducted within a natural intact oriental beech (Fagus orientalis) stands in district 2, located in the Experimental Forest of Gorgan university in the far east range of oriental beech stands in Hyrcanian forests, North of Iran. This region is situated between 36°41´ and 36°47´ N, and 54°20´ and 54°24´ E, covering a total area of about 1992 hectares with elevations ranging from 260 to 1960 m a.s.l. (Fig. 1).
The climate is Mediterranean with a mean monthly temperature ranging from 8.7 °C in February to 28.7 °C in July and means annual precipitation ~850 mm. Fagus orientalis followed by Carpinus betulus are the most frequent tree species contributing approximately 95% of the canopy cover in the study area. It also comprises other single trees including velvet maple (Acer velutinum Bioss.), cappadocian maple (Acer cappadocicum Gled.), Caucasian alder (Alnus subcordata C.A.May.), ironwood (Parrotia persica C.A.Mey), date palm (Diospyrus lotus L.), Broad-leaved lime (Tilia rubra Dc.) and wych elm (Ulmus glabra Huds.).
After an extensive reconnaissance in forest, we selected an area ~190 ha (parts of parcels 24 to 29) in closely pure F. orientalis vegetation zone with 870 to 1130 m a.s.l. with northern aspect and slope of ≤ 70%. The study area has involved typical old-growth characteristics, including very large and old trees, heterogeneous vertical and horizontal structure at small spatial scales and abundant coarse woody debris. Bedrock throughout the selected area is dominated by the limestone and soil texture is loam to clay-loam, soil type is classified as a forest brown soil.
2.2 Field sampling
In this study, two types of gaps were studied. The "canopy gap" was the land surface area directly under the canopy opening. The "expanded gap" consisted of the canopy gap plus the adjacent area extending to the bases of canopy trees surrounding the canopy gap (Runkle, 1981). The definition of canopy gap was important for determining if an open area would be sampled. Once we concluded that an area was a gap, we identified the surrounding perimeter tree boles to determine the boundaries of the expanded gap. To be included as part of the expanded gap boundary, all perimeter trees had to be dominant. Additionally, the gap perimeter had to be contiguous; with the ‘‘canopy connection’’ and distance between perimeter tree crowns not to be greater than 3 m (Almquist et al., 2002). Canopy gaps were defined as openings in the forest canopy larger than 20 m2 caused by the mortality of a tree with a diameter > 20 cm at 1.30 m height (DBH). Dead trees less than 20 cm DBH were not large enough to create a canopy gap (Nagel et al., 2010; Nagel and Svoboda, 2008; Bottero et al., 2011). When gapfillers reached a height of 24.5 m, the gap was considered closed. Of course, DBH and height of 150 trees, regularly distributed in all size classes, were measured to obtain stand height curves.
We sampled 76 gaps by using line-intercept sampling method along 5 transects varying in length from 625 to 2000 m parallel to slope contours. Transects were separated by 50 m to ensure that large gaps are not sampled more than once. When transect intersected an expanded gap, the following additional measurements were recorded. 1) The total length of transects, as well as the distance along transects intercepted by the canopy and expanded gap. 2) For each gap, radii (distance and direction) from the approximate gap center to the edge of each tree crown and corresponding bole that defined the gap was measured to the area of canopy and expanded gap were derived by nonconvex polygonal method here after called as NOPO method. 3) In order to calculate the area of the canopy gap and expanded gap based on ellipse formula the length L (largest distance from gap edge to gap edge) and width W (largest distance perpendicular to the length) was measured to the area of both kind of gaps were assessed by elliptical method here after ELLIP as an alternative method in gap area was surveyed. 4) DBH and species of gap border canopy trees 5) When possible, the characteristics of gap-makers comprising: species, DBH (if not possible the DBH class was determined), fall direction, decay class based on morphological characteristics to estimate the time since death of each gap-maker (Table 1), mode of mortality (i.e., standing dead, uprooted and snapped). Also, snapped gap-makers were categorized into one of three classes (i.e., snapped alive, snapped dead, or snapped unknown) (Nagel and Svoboda, 2008).

Table 1: Decay class system for gap-makers (after Nagel et al., 2006)
Forest structure was studied by sampling 65 circular plots of 1000 m2 along transects with 100 m intervals. In each plot with slope ≤ 70%, the DBH of all live trees ≥ 7.5 cm was measured. Trees were additionally placed into a pole-sized (DBH 7.5-20 cm) or canopy (DBH > 20 cm) height class. We used these data to calculate species importance values (IV) by averaging relative abundance, relative basal area, and relative frequency (max. IV = 100) using the technique outlined in Smith (1996).
2.3 Data analysis
The canopy and the expanded canopy gap area were determined using following calculation methods: 1) by fitting width (W) and length (L) dimensions to the formula for an ellipse (Equa. 1). 2) The area and perimeter of each gap were computed using a graphic editor computer program (AutoCAD ver. 2011) by the measured distance and direction from the center of each gap to the edge of each tree crown or corresponding bole for canopy and expanded gap, respectively (Nonconvex polygon). The area of canopy and expanded gap were classified in 100 and 200 m2 classes, respectively and the mid values of each class were showed in results.

In sampling with line transects, large gaps may be over-represented. Since the relative probability that a gap is crossed by the transect line is approximately equal to the gap diameter, the square root of the area of every gap, which is proportional to the diameter of a circular gap was taken based on Runkle (1982). As a result, gap size needs to be taken into account when estimating the gap fraction under the line-intersect sampling (LIS) approach. In addition, gap geometry also influences the estimation of gap fraction (Battles et al., 1996). Therefore, we used corrections to the standard LIS approach suggested by Battles and Fahey (1996), which takes both gap size and gap shape into account as Equa. 2.
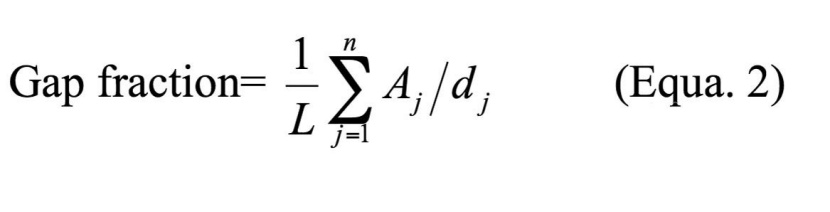
Where L is the transect length, Aj is the area of the jth gap encountered, and the dj is its effective diameter. The effective diameter (d) takes gap shape and size into account when estimating the probability of a line intersecting the gap and is equal to P/π, where P is the perimeter of the small convex cover of the object. P is simply equal to the actual perimeter of convex shapes, such as gaps shaped like ellipses. However, gaps measured in the field often have complicated shapes, form non-convex polygons, so P was used to calculate d (Battles et al. 1996). We used the expanded gap area to define inclusion for the gap fraction calculation. In other words, if the transect intersected only the expanded gap, we still included the canopy gap area in the calculation of gap fraction, but used the d value of the expanded gap to correct for the intersection probability. To describe the actual size distribution of gaps in the field, we also corrected for the inherent bias of oversampling large gaps with line transects following the suggestions of Battles et al. (1996).
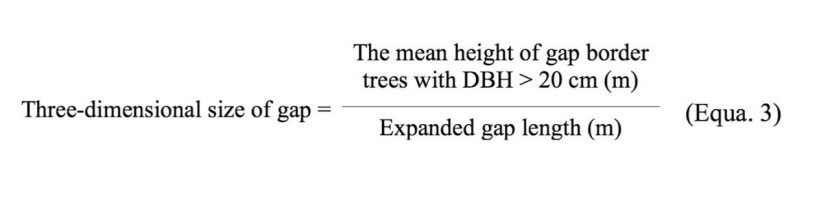
Whereas multiple-phase gaps may be formed due to different disturbance events, we used gap-maker tree decay classes and fall direction of tree simultaneously to estimate the number of disturbances in such gaps. We estimated the three-dimentional size of the expanded gaps based on the following formula and then categorized them based on Zhu et al (2005) classification’s criteria (Equa. 3).
The Pearson’s correlation coefficient was used for investigation of the linear relationship between paired data. The distribution pattern of the gap-maker’s state of decay and DBH classes was tested by Chi-square test. The normality and homogeneity of variance of quantitative data were investigated by Kolmogorov-Smirnov and Levene’s tests, respectively. We determined the differences of gap size as well as gap fraction in two groups (between NOPO and ELLIP methods)by dependent and independent t-test.
3. Results
3. 1 Stand structure
Mean density, volume and basal area of the investigated stands per hectare were 193.7 stems, 504.9 m3, and 35.8 m2, respectively and Fagus orientalis with an IV= 84.1 was the most important tree in the canopy layer of the forest. Other hardwood species such as Carpinus betulus, Acer velutinum, Parrotia persica and Diospyrus lotus were frequently associated with Fagus (Table 2). F. orientalis such as other tree species exhibited a reverse J-shape diameter distribution (Fig. 2).
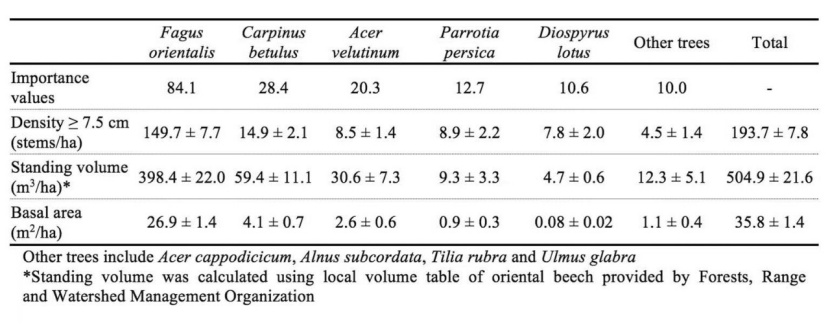
Table 2: Species importance values (IV = (Relative density + Rel. basal area + Rel. frequency)/3) and summary (mean±std) stand data in the site study
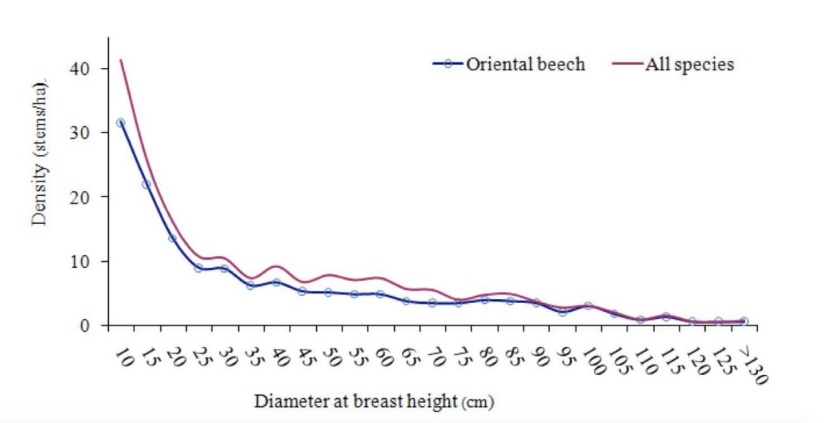
Fig. 2: Size distributions of Fagus orientalis and all trees (DBH ≥ 7.5 cm) in the site study
3.2 Gap characteristics and gap fraction
Seventy-six gaps occurred along the all transects with the 7.175 km length. The mean canopy gap fraction was 8.4% and 10.8% based on the gap area calculation method for nonconvex polygonal (NOPO) method and elliptical shape (ELLIP method), respectively. Gap fraction assessment were significantly different in two methods (t4 = -11.753, P < 0.01). Mean expanded gap fraction calculation from ELLIP method (22.0%) was greater than that from NOPO method (18.1%) in this region (t4 = -12.051, P < 0.01).
Gaps were highly variable in size; mean canopy gap size was 159 m2 and 208.7 m2 for NOPO and ELLIP methods, respectively. Mean expanded gap size was 515.7 m2 and 626 m2 for NOPO and ELLIP method, respectively over all five transects for two methods, respectively (Table 3).
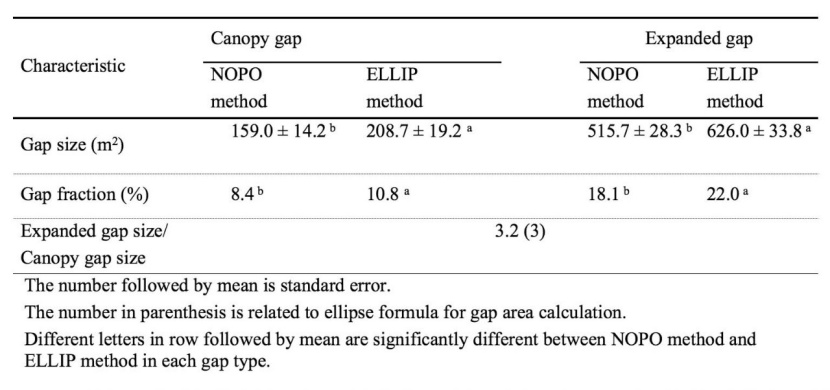
Table 3: Summary statics (Means) of canopy and expanded gap characteristics in an old-growth oriental beech forest in the site study
Canopy and expanded gap sizes, exhibited by ELLIP method was greater than that by NOPO method (t75 = -7.229, P < 0.01; t75 = -12.463, P < 0.01). Total area of canopy gap and expanded gap obtained from ELLIP method was 28% and 21% greater than those from NOPO method, respectively. The mean expanded gap/canopy gap was 3.2 for NOPO method that was about 6.25% greater than that for ELLIP method (3) (Table 3).
There were no significant difference between frequency distribution of canopy gap size in two methods of NOPO and ELLIP (Fig. 3), (Z = 0.649, p = 0.749). The size classes' distribution of expanded gaps was approximately normal in two methods. The maximum frequency of expanded gap was occurred on the 500 m2 size class of each two methods (Fig. 3) and no significance differences was detected between two methods (Z = 1.054, p = 0.216). The quadratic and linear equation obtained between the calculated gaps sizes of two methods for canopy (SC.non. = -1.2603 + 0.0002 S2C.elip + 0.8262 SC.elip. R2 = 0.92) and expanded gaps (SE.non. = 5.4665 + 0.8152 SE.elip. R2 = 0.96), respectively. Where SC.non. and SC.elip. are the area of canopy gap based on nonconvex polygonal and elliptical methods and SE.non. and SE.elip. are the area of the expanded gap based on nonconvex polygonal and elliptical methods, respectively.
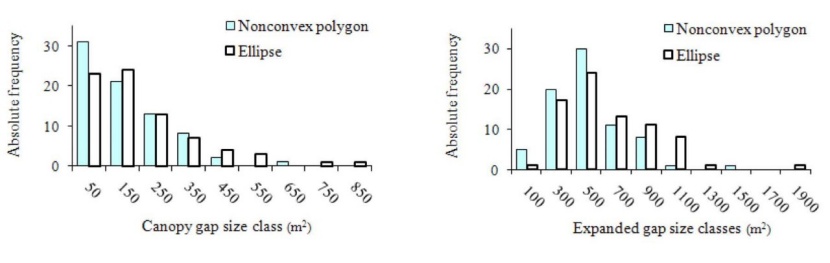
Fig. 3: Frequency distribution of canopy and expanded gap sizes in an old-growth oriental beech forest in the site study
Most of the canopy gaps (68.4%) were smaller than 200 m2, whereas only 1.4% of the gaps were larger than 500 m2 that comprised 5.8% of the total gap areas. In contrast, 41% of canopy gaps were smaller than 100 m2 that their proportion of the total gap areas was nearly 2.5 fold of gaps > 500 m2. However, the proportion of gaps with middle sizes (100-500 m2) were considerable (5 fold of gaps < 100 m2) (Table 4). Contrary to the canopy gaps, the expanded gaps were more frequent in middle size class (200-1000 m2) and nearly 90% of number and area of total gaps were corresponded to this size class (Table 4).
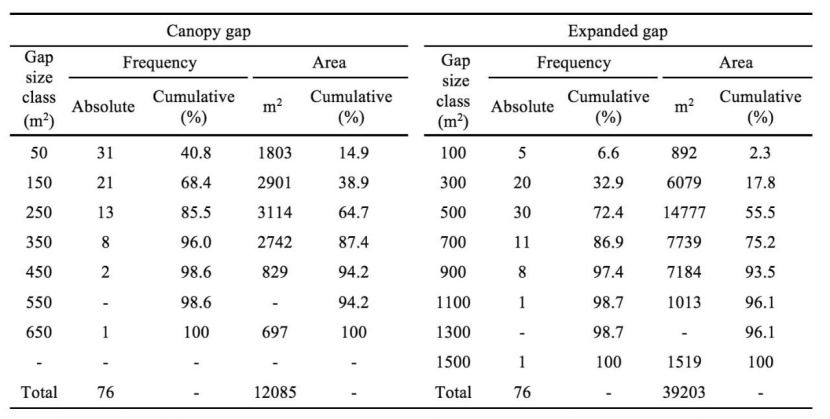
Table 4: Frequency and area distribution of canopy and expanded gaps in different size classes in the old-growth oriental beech forest in the site study
Our result showed as canopy gap size increased, the gap area-perimeter relationship differed markedly from circular gaps, indicating that gap geometry became more complex with gap size. The expanded gap also followed this pattern, but in lower sizes (area <450 m2) the complexity of expanded gaps was less than that of the canopy gaps (Fig. 4).
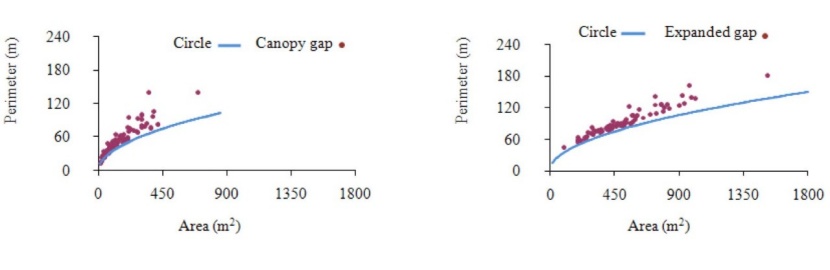
Fig. 4: Distribution of canopy and expanded gaps area-perimeter relationship in comparison with a circle shape in the old-growth oriental beech forest in the site study
While, frequency of expanded gaps was determined based on both size and three-dimensional classes (expanded gap length/mean height of gap border trees), the gaps < 400 m2 were classified as small gaps (ratio 0.5-1) and the gaps > 600 m2 (excluded of only one gap from 700 m2 size class) categorized as intermediate gap (Table 5). However, only gaps of 500 m2 size class (400-600 m2), were laid on each two three-dimensional classes. As a whole, most of the sampled gaps (nearly 62%) had a small size and no gap with large size (ratio > 2) was detected in the study area.
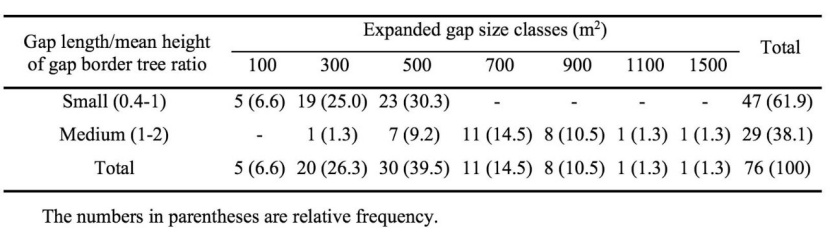
Table 5: Frequency (relative F.) of expanded gaps at three-dimensional size classes (after Zhu et al., 2015) in the old-growth oriental beech forest in the site study

Table 6: Frequency of gaps by number of disturbing events and of gap-maker tree in the old-growth oriental beech forest in the site study
3.3 Gap-maker characteristics
As a whole, from 76 recognized gaps, nearly half of them (47.4%) were formed by the death of one tree and the rest had two or more gap-makers. Although the maximum number of gap-makers per gap was 6 canopy trees, but such gaps rarely were seen in the study area. The use of decay classes of gap-maker trees to identifying the recurrence of disturbance events in each gap indicated that the most of gaps (56.6%) formed due to a single disturbing event. These gaps include of many gaps with multiple gap-makers (2 and 3); however, their portion was less than 10% of total gaps. We also showed that the mortality of 43% gap-makers was asynchronous, indicating of such gaps expanded after initial gap formation (Table 6). In spite of, several disturbance events had taken place in the study area, but many of single-phase gaps did not experience the recurrence of disturbance events (Table 7).
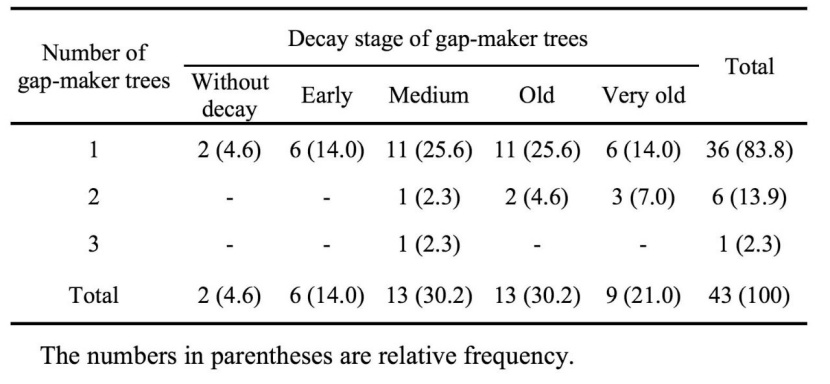
Table 7: Frequency of single-phase gaps by number of gap-maker tree and decay stage in the old-growth oriental beech forest in the site study
Generally, of the 139 existing gap-makers within 76 gaps, 78% were oriental beech and 10%, 2.8%, 2.8% and, 1.4% Carpinus betulus, Acer velutinum, Parrotia persica and Alnus subcordata, respectively; the remaining (4.4%) were considered unknown species because of their advanced decay state (Table 8). The analysis showed that the species composition of gap-makers was similar to that of gap border trees (χ24 = 3.131; P > 0.05) and so the forest canopy trees (χ24 = 4.014; P > 0.05). The canopy trees (trees > 20 cm) of the gap border tended to be larger in diameter compared to over-storey trees of forest stands (Z = -3.802; P < 0.01). This is while that nearly 20% of gap-makers were in the very old decay stage, and measuring of the diameter of them was not possible; therefore, gap-makers were classified in five categories (Fig. 5). The distribution pattern of gap-makers in these categories was differed form gap border trees (χ24 = 78.455; P < 0.01) and canopy trees of forest stands (χ24 = 120.078; P < 0.01). We could not identify the mode of mortality of 12.3% gap-maker trees; however, the dominant mode of gap-maker mortality was snapped-alive (59.7%). Standing dead gap-makers were present only in very small proportions (2.1%) in the study area (Table 8) while, the mortality of the half of the trees recognized as "snapped-unknown" and "unknown" was related to endogenous agents (pests and diseases). So, the role of such agents is very poor in comparison with exogenous agents.
The decay classes differed to such an extent in their morphological characteristics that should cover a broad enough time period to be representative of asynchronous gap-maker mortality. Identification of decay stage of a few gap-maker trees (2.2%) was impossible; nevertheless, the gap-makers were represented at all stages and mostly in old class (Table 8). The distribution pattern of gap-makers in different decay stages was non-uniform (χ24 = 47.084; P < 0.01).
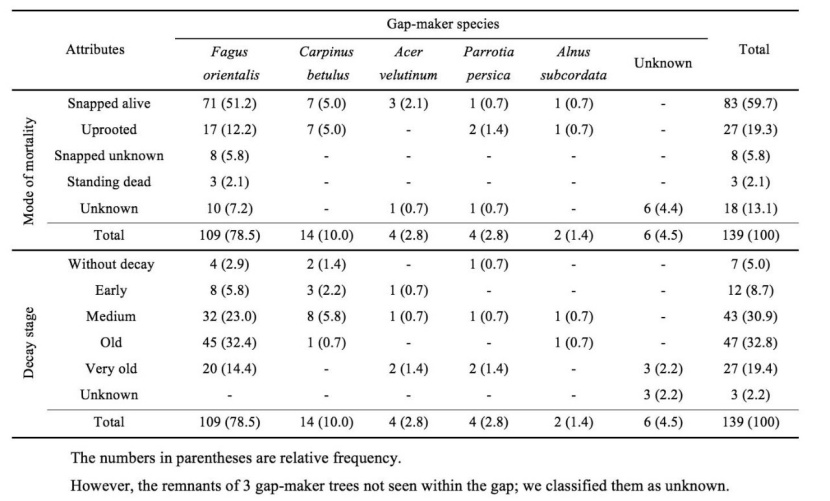
Table 8: Frequency of gap-maker trees by the mode of mortality, decay stage and species in the old-growth oriental beech forest in the site study

Fig. 5: Distribution of forest canopy tree, gap border tree and gap-makers in DBH size classes in an old-growth oriental beech forest in the site study
4. Discussion
4.1 Gap fraction
Mean canopy gap fraction in our study area was 8.4% that is almost similar (9.3%) to old-growth F. orietalis dominated forests in middle Hyrcanian forests of northern Iran (Sefidi et al., 2011), but it was almost twice that of other old-growth forests of eastern Hyrcanian (Amiri et al., 2015). The differences between reported gap fractions could be resulted from sampling methods and the criteria used to define gaps and from the gap areas calculation in these studies. In reality, in two mentioned studies, the full calipering method was used to study of gap fraction and the gap shape was assumed ellipse.
The overestimating of the ellipse equation for calculating of gap area was shown by Battles et al. (1996) and the present study (with a high intensity, about 31%). This issue caused the gap fraction of 8.4% exceeds to 10.8% in our study. In addition, surveying a large number of gaps and a large area improved the accuracy of the method (Yamamoto et al., 2011). It is noticed that the previous study in Hyrcanian forests had a limited area [(e.g. 3 separate blocks 25 ha in research of Sefidi et al. (2011) and one block 16 ha in research of Amiri et al. (2015)] and was carried out in mixed beech old-growth stands. Whereas, in our research the pure stands and integrated area that is larger than the sites of other studies was selected for this research.
Many factors contribute to gap regime heterogeneity within and among forest stands, including wind and fire patterns, stand age, stand composition, and site hydrology (Almquist et al., 2002). Yamamoto and Nishimura (1999) showed that the gap fraction in old-growth stands was greater than that of in the stands with initial developmental stages. The mean gap fraction in the present research was 8.4% in different elevation of the site study. The dendroecological analysis in an old-growth European beech forest in Bosnia and Herzegovina (Bottero et al., 2011) suggested that gap formation varied in time. Meanwhile, crown encroachment of trees surrounding newly formed gaps reduced in size, a fact that might be caused an error in area-based analyses of the rate of gap formation and disturbance intensity (Frelich and Martin, 1998). Lu et al. (2015) pointed out that larger gaps had a higher closure rate (39 m2/year) than middle gaps (25 m2/year) and small gaps (11 m2/year). According to the quadratic equations, the lateral growth could last 11, 13 and 16 years for small, middle and large gaps with a remaining size of 12, 69 and 223 m2, respectively.
The studies on the gap disturbance regime of intact beech forests in Europe have also varied results on the gap fraction. The gap fraction in oak-beech forest of Romania, 12.8% (Petritan et al., 2013), beech-fir forest of Slovakia, 11.3% and 15-16% (Kucbel et al., 2010; Drösser and Van Lüpke, 2005), mixed beech and coniferous forests of Bosnia and Herzegovina, 14% and 19% (Nagel and Svoboda, 2008; Bottero et al., 2011), pure beech forests of Slovenia, 5.6% (Zeibig et al., 2005), and mixed beech and coniferous forests of Czech, 9%-11% (Kenderes et al., 2008) was reported. Reports from other beech forests indicated that the gap fraction is also dissimilar, for example, this attributes in American mixed-beech forests was 9.5% and 17.7% (Runkle, 1982; Weskittel and Hix, 2003) and in beech forests of Japan 1.7% to 20% in stands with different age classes (Yamamoto and Nishimura, 1999). In addition to the previously mentioned factors, further factors may affect the gap fraction in mixed beech-coniferous forests that can be referred to the proportion of such species in the composition of canopy trees, as conifers have less capacity for lateral crown expansion compared with broadleaves (Nagel and Svoboda, 2008).
4.2 Gap characteristics
A negative exponential form, 41% and 58% of the gaps belonged to the small (<100 m2) and medium (100-500 m2) gaps and more than half less than 120.6 m2. However, nearly 79% of whole gaps area allocated to medium-size gaps indicating the prominent influence of such gaps on forest dynamics in comparison with small gaps. At the same time, small gaps are more likely to be closed by the lateral crown growth of gap border trees. Like Nagel and Svoboda (2008)’ finding, a high mean expanded gap/canopy gap ratio of small gaps in our study (6.5) confirms the importance of this issue (). The maximum canopy gap size in oriental beech forests of Iran often have been reported from 622 to 1250 m2 (Delfan-Abazari et al., 2004; Amiri et al., 2015); however, the all of these researches support the role of gaps < 500 m2, especially medium gaps in the dynamics of forest stands, whereas the large gaps (> 1000 m2) are known as a rare occurrence in these forests. The European beech forests studies show that the frequency of canopy gap sizes are followed by a negative exponential form (Nagel and Svoboda, 2008; Bottero et al., 2011) and log-normal (Drösser and Van Lüpke, 2005). The gaps < 100 m2 in these studies are more frequent (61% to 70%) than those in Hyrcanian beech forests of Iran. This issue can be resulted from the minimum area which was explained for 1) gap, 2) the age structure of gap 3) the presence of coniferous trees with a relatively smaller crown compared to broad-leaved trees in the composition of stands. Nevertheless, in European beech forests, the role of medium and large gaps in forest dynamics is by far more than of small gaps (Zeibig et al., 2005; Nagel and Svoboda, 2008; Kucbel et al., 2010).
The irregularity of gap shape that is related to the architecture of tree crown, the various responses of trees established in the gap border to the openings which were occurred in the forest canopy and the age, size and expansion of gap, was seen in our study site. As gap size increased, the shape of gaps differed markedly from circular diagram that is contrary with the results of Sefidi et al. (2011) but similar to those of Lertzman and Krebs (1991), Battles and Fahey (1996) and Nagel and Svoboda (2008). In general, irregular narrow gaps will receive far less photosynthetically active radiation at ground level than circular gaps of the same size (Muscolo et al., 2015). Meanwhile, gaps with a large perimeter to area ratio are more likely to be closed by lateral crown expansion of gap border trees (Battles et al. 1996).
4.3 Gap makers
The individual gaps with 2-6 gap-maker trees included more than half of the gaps in our study area. Some of the gaps with 2 and 3 gap-makers were formed due to the concurrent death of trees resulting from a single disturbance event, and about 43% of the gaps, experienced 1, 2 or 3 subsequent disturbing events after their initial formation. This issue revealed that such individual gaps formed from the expansion of smaller openings and no evidence existed that implicating smaller gaps were joined together at the length of time. Of course, it can be stated that age mortality, particularly beech tree that is not known for being very long lived, may be resulted in higher susceptibility of big tress towards wind throw, due to stem rotting and weaker root structure in the senescence phase.
There is not any report regarding multiple-tree gaps showing such gaps resulted from a single event or several disturbing events are caused by the individual gap formation in the Hyrcanian Forest Regions. In various researches the maximum number of gap-maker was reported from 4 -7 trees within an individual gap (Delfan-Abazari et al., 2004; Sefidi et al., 2011; Amiri et al., 2015). As a whole, in the mentioned studies as well as in the present study, such individual gaps were rare and often contained less than 4 gap-maker trees. The majority of gaps in the mixed beech-fir forests of Bosnia and Herzegovina were multiple-phase (Nagel and Svoboda, 2008; Bottero et al., 2011). Conversely, the most of the gaps in beech forests of Czech formed from the simultaneous death of 1-3 trees at the same place (Kenderes et al., 2008). Gap expansion due to chronic wind stress has also been reported in subalpine Picea-Abies forest in the northeastern United States (Worrall et al., 2005), as well as in a tropical semi-deciduous forest in southeastern Brazil (De Lima et al., 2008). Therefore, the gap expansion at the length of time could not be received as a prevalent phenomenon between all of the multiple-tree gaps. In spite of several disturbance events happened in the study area, many of single-phase gaps did not experience the recurrence of disturbing events.
This reality can be indicated that other reasons probably interfered in the expansion of gaps. Bottero et al. (2011) demonstrated that the endogenous agents, most likely related to wind and snow, act mainly as secondary agents in breaking weakened trees and in expanding previously established gaps. Likewise, a considerable number of gap-maker trees interfering with initial forming or in subsequent expansions of the individual gap had structural defects. Hardwood trees exposed by gap initiation and expansion will suffer crown damage due to the wind and deteriorate as decay fungi invade broken branches and wounds (Worrall et al., 2005). Young and Hubbell (1991) showed that the crown of most gap border trees was strongly asymmetrical into their adjoining gaps, and had a strong and significant tendency to fall on these heavy sides. However, the expansion of multiple tree gaps due to the occurrence of several disturbing events could be received as an important process in gap formation in our study area.
To some extent, the small and the medium gaps (mainly resulted from the expansion of smaller gaps) in our study indicate that periodic low-intensity disturbance events play an important role in forest dynamics. Conversely, Drösser and Van Lüpke (2005) and Nagel and Svoboda (2008) recorded several gaps > 0.4 ha with > 50 gap-makers in Slovakia and Bosnia and Herzegovina forests. This shows that the periodic disturbance events with intermediate intensity have an important role in these forests. Furthermore, Splechtna and Gratzer (2005) suggest the discontinuous age class distributions and the occurrence of peaks in the disturbance chronology of Alpine forests which can indicate the occurrence of intermediate disturbances (400-5000 m2) or possibly synchronized fine-scale disturbances and their importance for stand dynamics. Firm et al. (2009) suggest that periodic, intermediate severity wind disturbances may have had an important influence on pre-settlement forest dynamics in mixed mountain forests of the Alps. While Trotsiuk et al. (2012) states that forest dynamics are driven by periodic small disturbances, and larger events rarely occur in the beech forest. The high percentage of rotten trees in the upper canopy indicates that individual trees are prone to wind-breakage, which promotes these small-scale dynamics. Of course, the various pathogens can have make gap-makers more susceptible to wind or snow related mortality (Nagel and Svoboda, 2008). The stem breakage in the different height of trees and tree uprooting by wind was the dominant mode of gap-maker mortality in the study area. In spite of, we observed the stem structural defects in a considerable number of such gap-makers, but the exogenous agents, especially chronic winds probably had a prominent role in initial formation and expansion of gaps.
We were not able to identify the type of species of all gap-maker trees; nonetheless the oriental beech was the most frequent gap-maker among different species in the study site. Species composition of gap-makers were similar to both forests stand and gap-border canopy trees (trees with DBH >20 cm), indicating that the disturbances resulting from exogenous agents are more important in forest dynamics in comparison with endogenous agents; otherwise, the frequency of some species exposed to pests or disease should be more on the species composition of gap-maker trees. Ott et al. (2002) also found the same conclusion in their study of gaps in temperate rainforests of southeast Alaska. Meanwhile, gap etiology appeared to be rarely restricted in time to a single event, and was often complex, with multiple agents occurring in the same gap and on the same tree (Worrall and Harington, 1988). On the other hand, large gap-makers (trees with DBH >82.5 cm) were significantly more frequent than those trees that are present in the canopy layer of surrounding gaps and forest stands. The studies of old-growth forests in the other places of the world have had similar results (Ott et al., 2002; Worrall et al., 2005). Thus, probably the larger trees in the forest stands, more than other trees are threatened by the exogenous disturbance events.
5. Conclusion
The research findings are on the strength of the data which truly comprise a major part of the residual legacies on the surface of old-growth beech stands; those that the result of disturbance events has happened during recent decades in the study site. Data from elevations ranging from 900 – 1100 m indicate that canopy gaps are not static but some of them expand over time because mortality of gap-border trees and of gap expansion is more frequent than that of gap initiation at recent decades. The gap area calculating method influences the gap fraction but it can be estimated the actual area of gap obtained by elliptical method with a high accuracy. Meanwhile, the disturbance regime is described by the formation of small gaps as well as medium canopy gaps created from expanse of the former small gaps. Furthermore, most gaps were formed by snapped and uprooted trees, indicating the chronic winds had a key role in gap formation and expansion processes. The results suggest that some of single-tree gaps were expanded over the time; this is an important issue and should be studied in detail. It can be also stated that findings of the current study can be useful for implementation of close-to-nature silviculture in the Hyrcanean oriental beech stands. Likewise, the observed canopy gap characteristics can be guidance for forest managers towards practices that ensure forest sustainability.
References
Almquist B.E., Jack S.B., Messina M.G., 2002. Variation of the Treefall Gap Regime in a Bottomland Hardwood Forest Relationships with Microtopography, Forest Ecology and Management, 157: 155-163.
Amiri M., Rahmani R., Sagheb-Talebi KH., 2015. Canopy Gaps Characteristics and Structural Dynamics in a Natural Unmanaged Oriental Beech (Fagus orientalis Lipsky) Stand in the North of Iran, Caspian Journal of Environmental Sciences, 13(3): 259-274.
Attiwill PM., 1994. The Disturbance of Forest Ecosystems: the Ecological Basis for Conservative Management. Forest Ecology and Management, 63:247−300.
Battles J.J., Dushoff J.G., and Fahey T.J., 1996. Line Intersect Sampling of Forest Canopy Gaps. Forest Science 42: 131-138.
Bottero A., Garbarino M., Dukic V., Govedar Z., Lingua E., Nagel T.A., Motta R., 2011. Gap-Phase Dynamics in the Old-Growth Forest of Lom, Bosnia and Herzegovina, Silva Fennica, 45(5): 875-887.
Brang P., 2005. Virgin Forests as a Knowledge Source for Central European Silviculture: Reality or Myth? For. Snow Landsc. Res. 79: 19-32.
Brokaw N.V.L., 1985. Gap-phase Regeneration in a Tropical Forest, Ecology, 66: 682-687.
Canham C.D., 1988. An Index for Understory Light Levels in and Around Canopy Gaps, Ecology, 69: 1634-1638.
Delfan Abazari B., Sagheb-Talebi Kh., Namiranian M., 2004. Regeneration Gaps and Quantitative Characteristics of Seedlings in Different Development Stages of Undisturbed Beech Stands (Kelardasht, Northern Iran), Iranian Journal of Forest and Poplar Research,. 12(2): 302-306 [in Persian].
De Lima R.A.F., Martini AMZ, Gandolfi S, Rodrigues RR. 2008. Repeated Disturbances and Canopy Disturbance Regime in a Tropical Semi-Deciduous Forest. Journal of Tropical Ecology, 24: 85−93.
De Lima R.A.F., Prado P.I., Martini A.M.Z., Fonseca L.J., Gandolfi S., Rodrigues R.R., 2013. Improving Methods in Gap Ecology: Revisiting Size and Shape Distributions Using a Model Selection Approach, Journal of Vegetation Science, 24(3): 484-495.
Drösser L., Van Lüpke B., 2005. Canopy Gaps in Two Virgin Beech Forest Reserves in Slovakia, Journal of Forest Science, 51 (10): 446-457.
Firm d., Nagel T.A., Diaci J., 2009. Disturbance History and Dynamics of an Old-growth Mixed Species Mountain Forest in the Slovenian Alps, Forest Ecology and Management 257: 1893-1901.
Franklin J.F., Mitchell R.J., and Palik B.J., 2007. Natural Disturbance and Stand Development Principles for Ecological Forestry, USDA For. Ser. Northern Research Station Gen, Tech. Rep, NRS-19, 44p.
Frelich L.E., Martin G.L., 1988. Effects of Crown Expansion into Gaps on Evaluation of Disturbance Intensity in Northern Hardwood Forests, Forest Science, 34(2): 530-536.
Frelich L.E., Reich P.B., 2003. Perspectives on Development of Definitions and Values Related to Old-growth Forests, Environmental Reviews, 11: 9-22.
Gagnon J.L., Jokela E.J., Moser W.K., Huber D.A., 2004. Characteristics of Gaps and Natural Regeneration in Mature Longleaf Pine Flatwoods Ecosystems. Forest Ecology and Management, 187: 373−380.
Gray A.N., Spies T.A., Easter M.J., 2002. Microclimate and Soil Moisture Responses to Gap Formation in Coastal Douglas-fir Forests. Canadian Journal of Forest Research, 32: 332-343.
Kohm K.A., Franklin J.F., 1997. Creating a Forestry for the 21st Century: The Science of Ecosystem Management, Washington, DC: Island Press, 491 p.
Kenderes, K., Kamil, K., Vrška, T., Standovár, T., 2008. Natural Gap Dynamics in a Central European Mixed Beech-Spruce-Fir Old Growth Forest, Ecoscience, 16 (1):39-47.
Kucbel S., Jaloviar P., Saniga M., Vencurik J., Klmas V., 2010. Canopy Gap in Old-growth Fir-Beech Forest Remnant of Western Carpathians, European Journal of Forest Research, 129: 249-259.
Lertzman K.P., Krebs C.J., 1991. Gap-Phase Structure of a Sub-Alpine Old-Growth Forest, Can. J. For. Res., 21: 1730-1741.
Lundquist JE, Beatty JS. 2002. A Method for Characterizing and Mimicking Forest Canopy Gaps Caused by Different Disturbances. Forest Science, 48: 582-594.
Mataji A., Babaie Kafaki A., Safaee H., Kiadaliri H., 2008. Spatial Pattern of Regeneration Gaps in Managed and Unmanaged Stands in Natural Beech (Fagus orientalis) Forests, Iranian Journal of Forest and Poplar Research, 16 (1): 149-157. [In Persian].
McCarthy J.W., 2001. Gap Dynamics of Forest Trees: A Review with Particular Attention to Boreal Forests. Environmental Review, 9: 1−59.
Midgley J.J., Cameron M.C., Bond W.J., 1995. Gap Characteristics and Replacement Patterns in the Knysna Forest, South Africa, Journal of Vegetation Science, 6: 29−36.
Muscolo A., Bagnato S., Sidari M., Mercurio R., 2015. A Review of the Roles of Forest Canopy Gaps, Journal of Forestry Research, 25(4): 725−736.
Nagel T.A., Diaci J., 2006. Intermediate Wind Disturbance in an Old-Growth Beech-Fir Forest in Southern Slovenia. Canadian Journal of Forest Research, 36: 629-638.
Nagel T.A., Svoboda M., 2008. Gap Disturbance Regime in an Old-Growth Fagus-Abies Forest in the Dinaric Mountains, Bosnia-Herzegovina, Canadian Journal of Forest Research, 38: 2728-2737.
Nagel T.A., Svoboda M., Rugani T., Diaci J., 2010. Gap Regeneration and Replacement Patterns in an Old-growth Fagus-Abies Forest of Bosnia-Herzegovina, Plant Ecology, 208: 307-318.
Ott R.A., Juday G.P., 2002. Canopy Gap Characteristics and Their Implications for Management in the Temperate Rainforests of Southeast Alaska, Forest Ecology and Management, 159: 271-291.
Petritan A.M., Nuske R.S., Petritan I.C., Tudose N.C., 2013. Gap Disturbance Patterns in an Old-growth Sessile Oak (Quercus petraea L.)-European Beech (Fagus sylvatica L.) Forest Remnant in the Carpathian Mountains, Romania, Forest Ecology and Management, 308: 67-75.
Pregitzer K.S., Goebel P.C., Wigley T.B., 2001. Evaluating Forestland Classification Schemes as Tools or the Maintenance of Biodiversity. Jornal of Forestry 99:33-40.
Rademacher C., Neuert C., Grundmann V., Wissel C., and Grimm V., 2004. Reconstructing Spatiotemporal Dynamics of Central European Natural Beech Forests: the Rule-Based Forest Model BEFORE, Forest Ecology and Management, 194: 349-368.
Runkle J.R., 1981. Gap Regeneration in Some Old-Growth Forests of the Eastern-United-States, Ecology, 62: 1041-1051.
Runkle J.R., 1982. Patterns of Disturbance in Some Old-Growth Mesic Forests of Eastern North America, Ecology, 63: 1533-1546.
Runkle J.R., 1990. Gap Dynamics in an Ohio Acer-Fagus Forest. Canadian Journal of Forest Research, 20: 632-641.
Sagheb-Talebi Kh., Schütz J.P., 2002. The Structure of Natural Oriental Beech (Fagus orientalis) Forests in the Caspian Region of Iran and the Potential for the Application of the Group Selection System, Forestry, 75(4): 465-472.
Sagheb-Talebi Kh., Delfan-Abazari B., Namiranian M., 2005. Regeneration Process in Natural Uneven-aged Caspian Beech Forests of Iran, Schweizerische Zeitschrift fuer Forstwesen 156, 12: 477-480.
Schliemann S.A., Bockheim J.G., 2011. Methods for Studying Treefall Gaps: A Review, Forest Ecology and Management, 261: 1143-1151.
Sefidi K., Marvie Mohadjer M.R., Reinhard M., Copenheaver C.A, 2011. Canopy Gaps and Regeneration in Old-growth Oriental Beech (Fagus orientalis Lipsky) Stands, Northern Iran, Forest Ecology and Management, 262: 1094-1099.
Seymour R.S., White A.S., and DeMaynadier, P.G. 2002. Natural Disturbance Regimes in Northeastern North America-Evaluating Silvicultural Systems Using Natural Scales and Frequencies, Forest Ecology and Management, 155: 357-367.
Smith D.M., 1993. Silvicultural Systems Program: British Columbia Ministry of Forests. Canada-British Columbia Forest Resource Development Agreement, B.C. Min. For. Res. Br., Victoria, B.C. 2899.
Smith R.L., 1996. Ecology and Field Biology, 5th ed. Harper-Collins, New York.
Splechtna B.E., Gratzer G., 2005. Natural Disturbances in Central European Forests: Approaches and Preliminary Results from Rothwald, Austria, Forest Snow and Landscape Research, 79(1/2): 57-67.
Trotsiuk V., Hobi M.L., Commarmot B., 2012. Age Structure and Disturbance Dynamics of the Relic Virgin Beech Forest Uholka (Ukrainian Carpathians), Forest Ecology and Management, 265: 181-190.
Ulanova N.G., 2000. The Effects of Wind Throw on Forests at Different Spatial Scales: A Review, Forest Ecology and Management, 135: 155-167.
Watt A.S., 1947. Pattern and Process in the Plant Community, Journal of Ecology, 35: 1-22.
Weiskittle A.R., Hix D.M., 2003. Canopy Gap Characteristics of an Oak-Beech-Maple Old-Growth Forest in Northeastern Ohio, Ohio Journal of Science, 103(4): 111-115.
White P.S., Pickett S.T.A., 1985. Natural Disturbance and Patch Dynamics: an Introduction. In: Pickett, S.T.A.; White, P.S., eds. The Ecology of Natural Disturbance and Patch Dynamics. New York: Academic Press: 3-13.
Whitmore TC. 1989. Canopy Gaps and the Two Major Groups of Forest Trees. Ecology, 70: 536−538.
Woods K.D., 2004. Intermediate Disturbance in a Late-Successional Hemlock Northern Hardwood Forest, Journal of Ecology, 92: 464−476.
Worrall J.J., Harrington T.C., 1988. Etiology of Canopy Gaps in Spruce-Fir Forests at Crawford Notch, New Hampshire, Canadian Journal of Forest Research, 18: 1463-1469.
Worrall J.J., Lee T.D., Harrington T.C., 2005. Forest Dynamics and Agents that Initiate and Expand Canopy Gaps in Picea-Abies Forests of Crawford Notch, New Hampshire, USA, Journal of Ecology, 93: 178-190.
Yamamoto S.I., Nishimura N., 1999. Canopy Gap Formation and Replacement Pattern of Major Tree Species among Developmental Stages of Beech (Fagus crenata) Stands, Japan, Plant Ecology, 140: 167-176.
Yamamoto S.I., Nishimura N., Torimaru T., Manabe T., Itaya A., Becek K., 2011. Comparison of Different Survey Methods for Assessing Gap Parameters on Old-growth Forests, Forest Ecology and Management, 262(5): 886-893.
Young T.P., Hubbell S.T., 1991. Crown Asymmetry Treefalls and Repeat Disturbance of Broad Leaved Forest Gaps, Ecology, 72(4): 1464-1471.
Zeibig A., Diaci J., Wanger S., 2005. Gap Disturbance Patterns of a Fagus sylvatica Virgin Forest Remnant in the Mountain Vegetation Belt of Slovenia, Forest Snow and Landscape Research, 79(1/2): 69-80.



