Schlüsselbegriffe: Acer velutinum, Alnus subcordata, Parrotia persica, Ingenieurbiologie, Hangstabilität, Grabenmethode
Abstract
The aim of this study was to investigate the spatial distribution and mechanical properties of tree roots of three pioneer species of the Hyrcanian forest including alder, maple, and Persian ironwood. As these species are frequent in disturbed areas and also occur on slopes, they can be used efficiently for bioengineering measures. We also assessed the effect of season on the mechanical properties of roots and the increased cohesion of soil supplied by the roots of the examined tree species. Six sample trees were selected randomly for each species, and two trenches were dug on the uphill and downhill sides of each sample tree. We next estimated the root area ratio (RAR), tensile force, tensile strength, and root reinforcement at each side of each sample tree. Our results showed that in shallower depths was RAR higher, and near profile walls exhibited significantly higher RAR than far walls. Downhill trenches likewise had significantly higher RAR values compared to uphill trenchers. On the other hand, the RAR values did not significantly differ between the three tree species. Tensile force and strength versus root diameter followed a positive and negative power law relationship, respectively. Tensile force was highest for Persian ironwood roots and tensile strength was significantly higher in winter compared to summer. Root reinforcement was significantly higher in winter, for Persian ironwood, at downhill slopes, and for shallow soil layers. The effect of the distance from the stem was only significant for Persian ironwood. Since little information regarding mechanical and biotechnical properties of these species have been available, this study now helps us to understand the root characteristics of important pioneer species needed to evaluate their contribution to root reinforcement and slope stability.
Zusammenfassung
Ziel der vorliegenden Studie war es, die räumliche Verteilung und die mechanischen Eigenschaften der Wurzeln von drei häufigen Pionierbaumarten Erle, Ahorn und Persischer Eisenholzbaum des Hyrkanischen Waldes zu untersuchen. Da diese drei Baumarten häufig auf gestörten Flächen und an Hängen vorkommen, könnten diese für ingenieurbiologische Massnahmen eingesetzt werden. Wir waren außerdem an saisonalen Muster in den mechanischen Eigenschaften der Wurzeln interessiert und an der Untersuchung der erhöhten Bodenkohäsion durch Wurzeln. Für jede Art wurden sechs Probenbäume zufällig ausgewählt und jeweils zwei Gräben an den hangober- und hangunterseitigen Seiten jedes Probenbaums gegraben. Auf jeder Seite der Probenbäume wurden dann Wurzelflächenverhältnis (RAR), Zugkraft, Zugfestigkeit und Wurzelverstärkung ermittelt. Unsere Ergebnisse zeigten, dass RAR in flachere Bodentiefen höher ist und dass die näheren Grabenwände signifikant höhere RAR-Werte aufweisen als die weiter entfernten Wände. Darüberhinaus hatten hangabwärts gelegene Gräben signifikant höhere RAR-Werte als hangaufwärts gelegene. Die RAR-Werte unterschieden sich jedoch nicht signifikant zwischen den Baumarten. Das Verhältnis zwischen Zugkraft und Festigkeit zum Wurzeldurchmesser hatte jeweils einem positiven bzw. negativen potenziellen Zusammenhang. Unter den untersuchten Baumarten war die Zugkraft am größten beim Persischen Eisenholzbaum und im Winter war die Zugfestigkeit signifikant höher als im Sommer. Der Abstand zum Stamm hatte nur für den Persischen Eisenholzbaum einen signifikanten Einfluss auf die untersuchten Wurzeleigenschaften. Da wir nur kaum Informationen zu den bodenmechanischen und biotechnischen Eigenschaften dieser Bäume haben, hilft unsere Studie nun bei der Bewertung der Rolle dieser Baumarten bei der Wurzelverstärkung und Hangstabilität.
Introduction
Forests play a significant role in reducing natural hazard risks such as landslides and erosion processes (Schmaltz et al., 2017). Slope instabilities are frequently recurring problems in the northern forests of Iran, known as the Hyrcanian forest, due to deep layers of cohesive soils and relatively high precipitation that lead to high moisture content in theses soils. Landslides occur in these forests and may lead to the destruction of infrastructures requiring considerable investments to repair them (Zare et al., 2013). Tree plantation may be one solution to prevent these destructive events and have been reported as a cost-effective and environmentally friendly measure for slope stabilization and erosion control. However, little is known on which plant species are best suited for stabilizing slopes. The selection of tree species for planting (reforestation or afforestation) in the context of soil protection against natural hazards is often chosen by default and made according to their adaptation to the environment, availability, financial considerations, and biological characteristics (Watson and Marden, 2004), rather than their biotechnical properties.
Using plants and trees with the purpose of slope stabilization has been considered widely as ‘Green Technology’ (Bordoloi et al., 2015). This technology is considered as a soft or green engineering method compared with traditional complicated approaches such as soil nails, anchors or meshes (Hytiris et al., 2015). Roots are known as important elements of soil stabilization which supply the plants with water and nutrients and provide stability for the plant itself (Heidarian et al., 2018). On the other hand, roots increase soil cohesion and therefore improve soil stability. Roots are well-known to enhance soil stability and control natural hazards like erosion, shallow landslides, and mass movements (Stokes et al., 2010). From a mechanical point of view, roots may anchor weaker soil layers to stronger layers below by providing additional resistance (Dumlao et al., 2015).
The most important factors used in soil reinforcement assessment are root density and root strength (Operstein and Frydman, 2000; Roering et al., 2003). Many biotic and abiotic factors such as site condition, topography, soil moisture content, soil type, tree species, climate and season influence root reinforcement (Schwarz et al., 2012; Vergani et al., 2016, 2017). The effects of environmental factors on the mechanical properties of roots have been frequently investigated already (Genet et al., 2005, 2011; Hales et al., 2013; Zhang et al., 2014; Yang et al., 2016). Another important environmental factors may be season, especially in temperate forest zones with a dominance of deciduous forests. Tensile strength has been reported to be higher in the leafless period than in the leafed period (Coppin and Richards, 1990; Karrenberg et al., 2003; Genet et al., 2005; Norris et al., 2008). Furthermore, significant differences have been reported for root properties in uphill versus downhill sides of the tree due to different magnitudes of load and stress. Root tensile strength in the uphill side has been reported to be higher compared to the downhill side (Gary and Sottier, 1996; Schmidt et al., 2001; Chiatante et al., 2003; Norris et al., 2008). In addition, a negative power relation between root diameter and tensile strength has been reported in many previous works (Genet et al., 2005; Nyambane and Mwea, 2011; Naghdi et al., 2013). Moreover, tensile force increases with increasing root diameter following a positive power relation (Nilaweera, 1994; Gray and Sotir, 1996; Bischetti et al., 2005; Ji et al., 2012; Vergani et al., 2012).
Tree plantation is a simple method with relatively low costs of establishment and maintenance and is used to reinforce slopes in many parts of the world (Hales and Miniat, 2017). Trees increase slope stability by root reinforcement and also increase soil cohesion (Watson and Marden, 2004; Genet et al., 2006, 2008; Hales and Miniat, 2017). Tree plantations help managers to increase the strength of soil, especially whenever root decay decreases soil cohesion (e.g., after clear cutting) or where there is no vegetation cover to protect soil from erosion, landslides and mass movements. Alnus subcordata (alder), Acer velutinum (maple) and Parrotia persica (Persian ironwood) are native and pioneer species in the Hyrcanian forest. They are light demanding and grow easily on slopes of the Hyrcanian forest, especially in forest gaps, even in relatively poor soils. However, there is a lack of information regarding mechanical and biotechnical properties of these trees in the same environmental condition so that they can be evaluated and compared in terms of soil protection and also can be assessed in terms of their contribution to root reinforcement and slope stability. Therefore, understanding the root characteristics of these tree species is necessary for such evaluations.
Concerning engineering applications, quantification of root reinforcement is a key parameter for several fields of application, including slope stability analysis (Vergani et al., 2012), erosion control measures (Giadrossich et al., 2016), and soil bioengineering technique design (Bischetti et al., 2010). Therefore, the aims of the current study were: i) to investigate the spatial distribution and mechanical properties of alder, maple, and Persian ironwood tree roots, ii) to assess the effect of season on the mechanical properties of the roots, and iii) to evaluate and compare the increased cohesion of soil supplied by alder, maple (as man-made plantations) and ironwood (natural stand) tree roots.
Material and methods
Study site
The study area is in Northern Iran within the northward slopes of Alborz Mountains, in the Hyrcanian forest. The Hyrcanian forest is categorized as natural deciduous broadleaved with uneven aged mixed stands. Regarding diameter distribution, the largest number of stems fall in the smallest diameter classes and the number of stems per hectare drops following a negative exponential function with increasing tree diameter. The regeneration usually depends on natural regeneration, and sometimes tree plantation is conducted in the disturbed stands. The studied area was located within the first district of the educational and experimental forest of the University of Tehran (namely the Patom district), in a part of compartment No. 112, situated between 36° 27'-36° 40' N and 51° 32'-51° 43' E., WGS84. The forest type in the compartment is Fageto-Carpinetum accompanied by Acer velutinum and Alnus glutinosa and consists of 154 stems per hectare and about 279 m3 standing volume per hectare. The tree species that existed in the compartment are Acer cappadocicum, Acer velutinum, Alnus glutinosa, Carpinus betulus, Diospyros lotus, Fagus orientalis, Parrotia persica, Quercus castaneifolia, Rhamnus frangula, Tilia begonifolia and Ulmus glabra. The mean annual precipitation in the area is about 1200 mm with October being the wettest month (average: 235 mm) and August the driest month (average: 42 mm). Precipitation in the area is falling mostly as rain and includes a cover of snow in winters. The mean annual temperature is 17 °C in the area with February and August as the coldest and warmest months, respectively (Deljouei et al., 2018). The site has mainly calcareous parent materials with many cracks and fine grain clay soil having low and high plasticity (CL and CH soil classes in USCS) as the most frequent soil classes. The results of Monitoring soil moisture content over one year showed that the mean and maximum soil moisture content was 20 and 36.8 %, respectively (gravimetric moisture content). Direct shear tests were used at these two moisture contents, to derive the soil strength parameters. This analysis resulted in values of c (soil cohesion) of 15.3 and 1.1 kPa and values of φ (soil friction angle) of 29 and 27 degrees for the moisture contents of 20 and 36.8 %, respectively. In some parts of the site along forest roads, shallow landslides were easily recognized. Most of the shallow landslides in this region occur after heavy rainfalls (Abdi, 2018). The critical plane of these landslides in many cases is less than 1 m. Therefore, vegetation can be useful to stabilize slopes. A mixed plantation of Alnus subcordata (alder), Acer velutinum (maple) and an adjacent natural stand of Parrotia persica (Persian ironwood) with an area of about 3 ha were selected for the current study. These species are native, pioneer and light demanding species of the temperate forest of Iran, which may be used efficiently for bioengineering measures in disturbed sites around forest roads (Abdi et al., 2010; Naghdi et al., 2013). The estimated volumes of alder, maple and ironwood in the compartment are 3.70, 4.69 and 1.90 % of total stand volume in the compartment, relatively. Six sample trees were selected randomly for each species and the mean (± standard error) diameters at breast height (DBH) of samples were 33.00±2.22, 30.50±1.56 and 30.80±2.40 cm for alder, maple, and ironwood, respectively. The mean (± standard error) heights of the sample trees were 20.94±1.99, 23.17±1.76 and 13.31±1.92 m for alder, maple and ironwood, respectively. The site is located on a relatively uniform northwest facing slope with 25 % gradient.
Root distribution
Six sample trees were selected randomly for each species and two trenches were dug at the uphill and downhill sides of each sample tree to assess the effect of slope on root properties. Therefore, the root distribution of alder, maple, and Persian ironwood was measured using 36 trenches and 72 profile walls setting up two profile walls on the two opposite sides of the trench, facing the tree. The trenches were dug down either to the maximum rooting depth (bedrock) or down to 1 m soil depth at the deepest, assuming that root density is very low below (Vergani et al., 2014). The nearest profile wall was established on the side closest to the tree (Figure 1) and the distant profile wall was established at the opposite side of the trench.
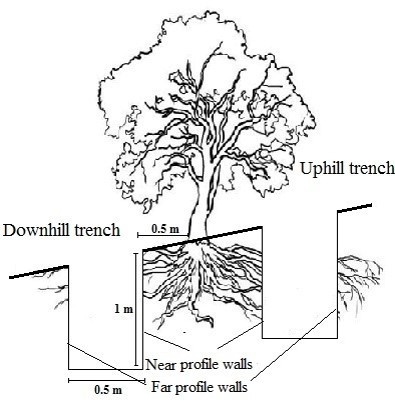
Figure 1: The schematic design of trenches relative to a sample tree and slope. / Abbildung 1: Der schematische Aufbau von Gräben relativ zu einem Probebaum und Hangneigung.
The trench dimensions were chosen to be 1 m length by 0.5 m width by 1 m depth, suggested to investigate the effect of slope on the tree root system (Ji et al., 2012). 10 cm layers were marked on the vertical profile walls using pins and string. Roots were counted and measured directly on the trench walls in June 2017 to estimate root density parameter (Schwarz et al., 2010; Vergani et al., 2014). The root diameters were measured by digital calipers (Abdi et al., 2014, 2019) and RAR was calculated as follows:

where di is the diameter of the i-th root (mm), Ar is the cross sectional area of roots (mm2), and As is the area of soil (mm2) for each 10 cm layer at each profile wall.
Root tensile strength
Root samples were collected with the diameter (including bark) ranging from 0.29 to 4.70 mm in two seasons, leafless (November 22, 2017) and leafed period (June 28, 2017), to investigate the effect of season on root strength. Root samples were collected randomly from soil by excavating small independent pits in the two seasons. The location of pits was at a distance of 30 cm from the stems, downslope side and adjacent to the trees down to a depth of 30 cm below the soil surface; and live roots were collected with sharp scissors. A 15 % alcohol solution (Burylo et al., 2011; Chiaradia et al., 2012) was only sprayed on the sample roots only instead of directly adding the solution to bags to avoid severe changes of moisture content. The specimens were transferred to the laboratory and root tensile strength was measured by a Universal Testing Machine (SANTAM Co./ SMT-5), equipped with 500 kg maximum-capacity load cell. Root samples with a length of 10 cm were positioned as vertical as possible with the load cell axis. The strain rate of 10 mm/min (Chiaradia et al., 2016; Vergani et al., 2016; Gilardelli et al., 2017) was applied until rupture occurred from the middle part of the roots (Genet et al., 2008; Ji et al., 2012). Root tensile strength was calculated at the moment of the maximum tensile force as follows (Preti, 2013):

where Tr (MPa) is the root tensile strength, Fmax is the maximum tensile force (N) and A0 is the root cross-sectional area (mm2).
Root reinforcement
The increased soil cohesion due to the presence of roots was estimated by a modified version of the Wu (1979) model based on the correction factor (k'') introduced by Preti (2006). The root reinforcement model of Wu is based on the Coulomb equation, in which soil shearing resistance is calculated based on cohesive and frictional forces as follows:

where τsr is the soil shearing resistance (kPa) which is measured with soil shearing tests, usually direct shear tests for slope stability, cv is the additional cohesion or increased shear strength due to the presence of roots (kPa), cs is the soil cohesion (kPa), σ is the normal stress at the shear plane (Pa), and ф is the soil friction angle (degree). Moreover, cs and ф are derived from direct shear tests and σ is the surcharge factor applied during shear tests.
When soil mass movement, landslides or erosion occurs, shear forces increase and roots crossing the slip surface are mobilized in tension. As a consequence, the additional cohesion cv is thus expressed as (Preti, 2013):

where tr is the mobilized root tensile strength per unit area of soil, k΄ is a factor taking into account the random orientation of roots with respect to failure plane, which is usually considered equal to 1.2 (ranging from 1.0 to 1.3: Waldron, 1977; Wu et al., 1979). The parameter k˝ reported to be 0.4 is an empirical correction factor introduced by Preti (2006) in order to correct the overestimation of the cv values in the Wu model. The mobilized root tensile strength is defined as:

where Tr is the average tensile strength per average root cross sectional area and RAR is the root area ratio.
Also, as there is a lack of information on the mechanical effects of fine roots compared to coarse roots and fine roots are important for primary production (Konopka, 2017), we calculated their reinforcement effect and compare it with coarse roots.
Data analysis
A Kolmogorov–Smirnov test was used to check the normality of the data before proceeding with analysis and where this assumption was violated (normality), data were transformed to ensure homogeneous residual variance and normality. The mean RAR differences among the species and profiles of the uphill and downhill trenches were tested for significance at the 0.05 probability level where the nested ANOVA indicated significant differences. As the roots were collected from the three species and two seasons, the variations of tensile force and tensile strength can be also assessed. To determine whether there were any differences in root strength in different tree species and seasons, the results were subjected to ANCOVA (the root diameter as the covariate, and all treatment have the same diameter ranges). Moreover, nested ANOVA was used to assess the effects of species, soil depth, season and distance from stem on root reinforcement. The data were analyzed using the SPSS (Ver. 20) statistical software.
Results
Root distribution
The results indicated that most of the roots that occurred on the profile walls were roots with diameters less than 10 mm. The number of roots decreased with increasing soil depth and most roots occurred at shallower depths (Figure 2).
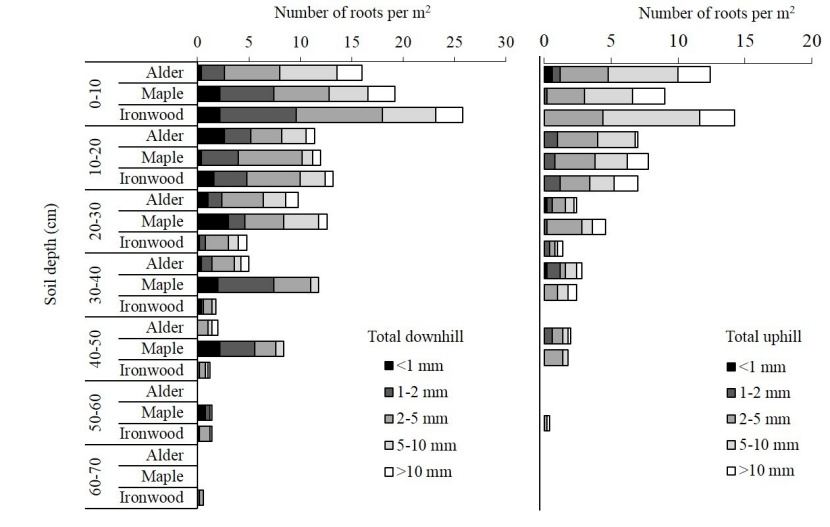
Figure 2: Total number of roots based on root diameter at different depths of the soil for both down and uphill of three different tree species. / Abbildung 2: Gesamtzahl der Wurzeln basierend auf der Größe des Wurzeldurchmessers in verschiedenen Bodentiefen, sowohl bergauf als auch bergab von drei verschiedenen Baumarten.
The maximum, minimum and mean values of RAR are shown in Table 1. According to the table, the ranges of the mean, max and min values of RAR were 0.15-2.72 %, 1.62-26.98 and 0.0005-0.14, respectively.
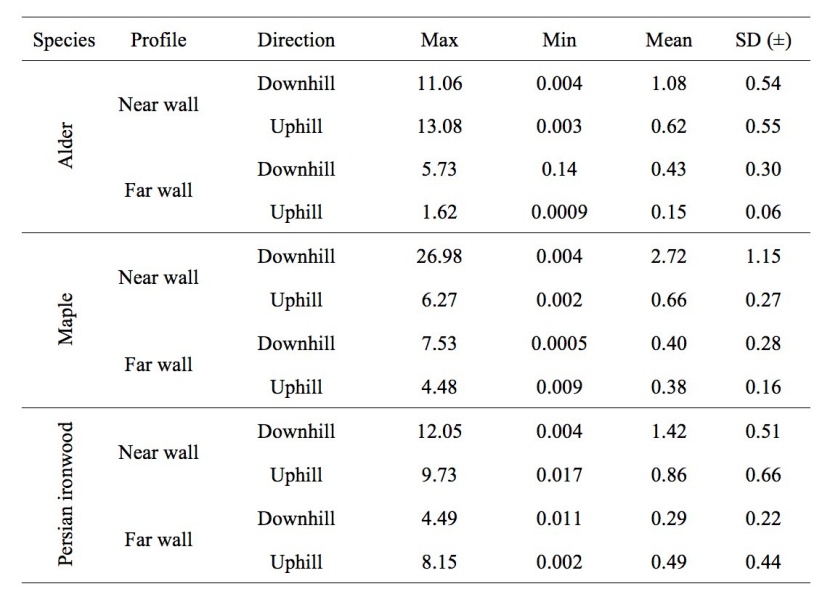
Table 1: Descriptive summary of the Root Area Ratio (RAR) values in %. / Tabelle 1: Zusammenfassung der Werte für Wurzel Flächenverhältnis (RAR) in %.
As Figure 3 shows, the RAR values of alder, maple, and Persian ironwood generally decreased with increasing soil depth. The results revealed that the maximum RAR values were located in the upper layer of soil (generally in the first 10 cm).
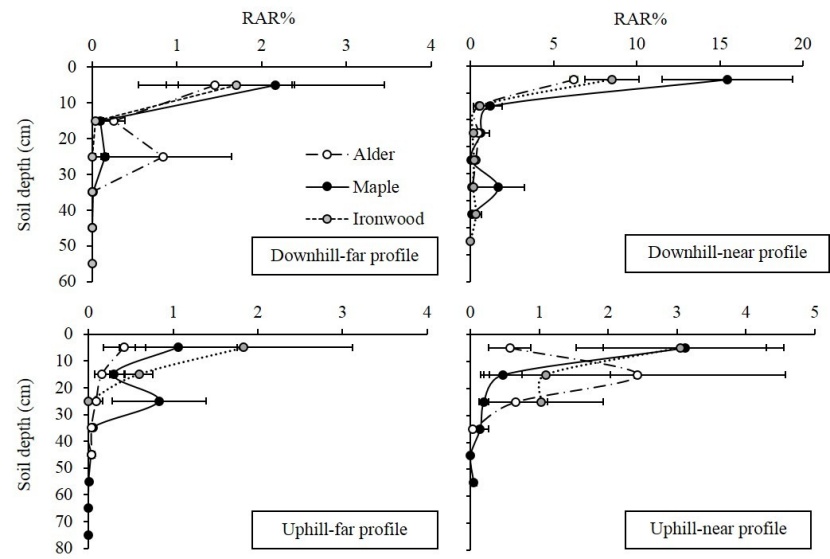
Figure 3: Mean values of Root Area Ratio (RAR) regarding species and different profile walls. / Abbildung 3: Mittelwerte des Wurzel Flächenverhältnis (RAR) für Arten und unterschiedliche Profilwände.
The results of the nested ANOVA showed that the effect of slope, distance from stem, and soil depth was significant on the RAR values in the three investigated species (p < 0.05). The results of the Duncan test showed that near profile walls on both sides of the tree had significantly higher RAR values than far profile walls (F=12.48, p < 0.05). Moreover, downhill trenches had significantly higher RAR values compared to uphill ones (F=6.98, p < 0.05). However, the RAR values were not significantly different between the tree species (F=2.98, p > 0.05).
Root strength
Descriptive statistics of the tested root sizes and their corresponding tensile force and strength values are shown in Table 2. As presented in Table 2, the number of valid tests was between 29 and 59 samples based on species and seasons. The root diameter ranged from 0.29 to 4.70 mm, and the mean root diameter for each species varied between 1.4 and 1.7 mm.
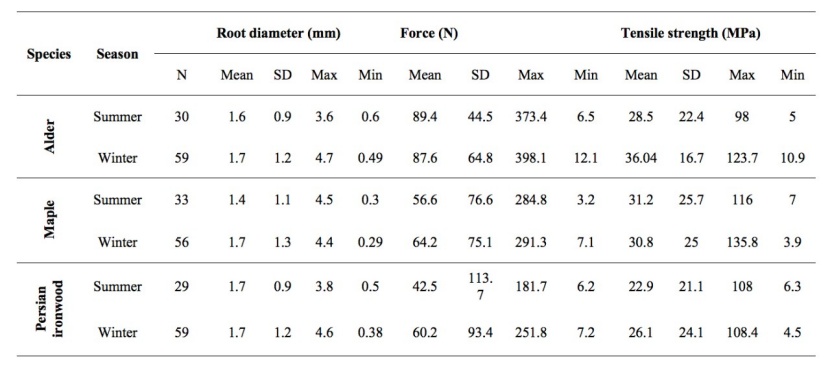
Table 2: Descriptive summary of the tested roots including diameter, force and strength. / Tabelle 2: Zusammenfassung der getesteten Wurzeln hinsichtlich Durchmesser, Kraft und Festigkeit.
The mean tensile force values for alder, maple and Persian ironwood for winter were 87.6, 64.2 and 60.2 N and the corresponding values for summer were 89.4, 56.6 and 42.5 N, respectively.
Concerning tensile strength, the mean tensile strength values of alder, maple and Persian ironwood for winter were 36.04, 30.8, and 26.1 MPa and the corresponding values for summer were 28.5, 31.2, and 22.9 MPa, respectively.
The relationships between tensile force and strength versus the root diameter were plotted and then the power laws were fitted to the data. The relationships between tensile force and the root diameter was followed by a positive power law. However, a negative power relation existed between tensile strength and the root diameter (Figures 4 and 5). The corresponding fitting curves are shown in Figures 4 and 5.
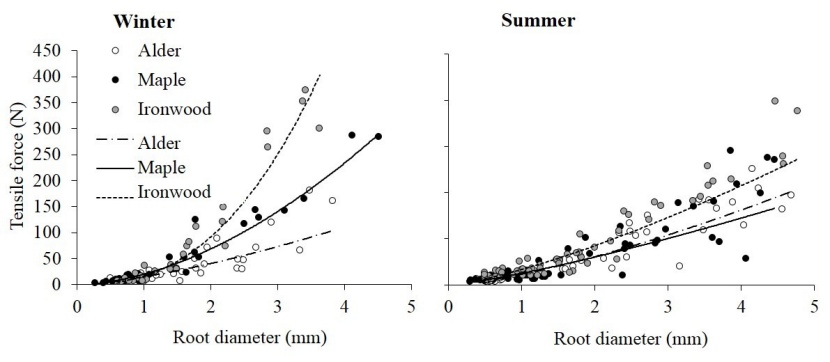
Figure 4: Tensile force versus root diameter in winter and summer. The lines show power regression curves fitted to the data of the three species. / Abbildung 4: Zugkraft versus Wurzeldurchmesser im Winter und Sommer. Die Linien zeigen Regressionskurven, die an die Daten der drei Arten angepasst sind.
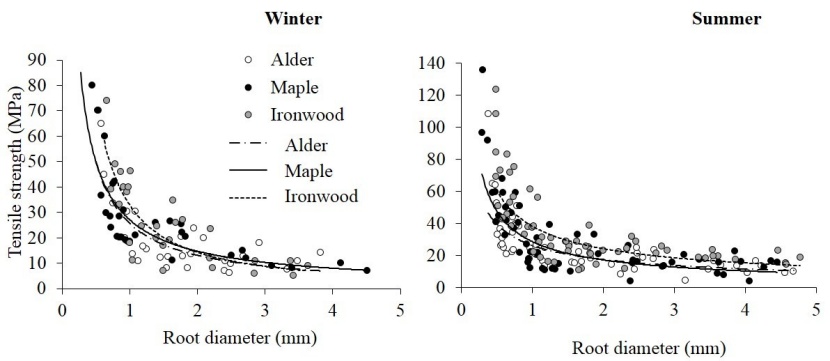
Figure 5: Tensile strength versus root diameter in winter and summer. The lines show power regression curves fitted to the data of the three species. / Abbildung 5: Zugfestigkeit gegen Wurzeldurchmesser im Winter und Sommer. Die Linien zeigen Regressionskurven, die an die Daten der drei Arten angepasst sind.
The results of ANCOVA (with the root diameter set as the covariate factor) revealed that there were no significant differences in tensile force between winter and summer (F=0.003, p > 0.05). However, there were significant differences among the species (F=22.10, p < 0.05). In this regard, Persian Ironwood had the highest strength, but alder and maple had no significant differences with each other. Furthermore, the root diameter as the covariate parameter was significant (F=981.18, p < 0.05).
Moreover, the results of ANCOVA revealed that tensile strength was significantly greater in winter than in summer (F = 4.71, p < 0.05) with regard to the root diameter as the covariate factor (F = 131.64, p < 0.001). No significant differences in tensile strength were found among the different species (F = 3.09, p > 0.05).
Root reinforcement
The mean root reinforcement values for alder, maple and Persian ironwood for winter were 0.51, 0.50, and 0.65 kPa and the corresponding values for summer were 0.33, 0.39, and 0.36 kPa, respectively. Comparing the role of fine (<2 mm) and coarse roots (>2 mm) in soil reinforcement showed that the mean effects of root classes are 22.42±4.46 and 77.58±4.48 % respectively.
The results of the nested ANOVA showed that root reinforcement between the uphill and downhill positions, between different distances from the tree and between various soil depths was significantly different in the three species (Table 3). Furthermore, root reinforcement between the three species was significantly different. The results of mean comparisons with the Duncan test are shown in Table 3.
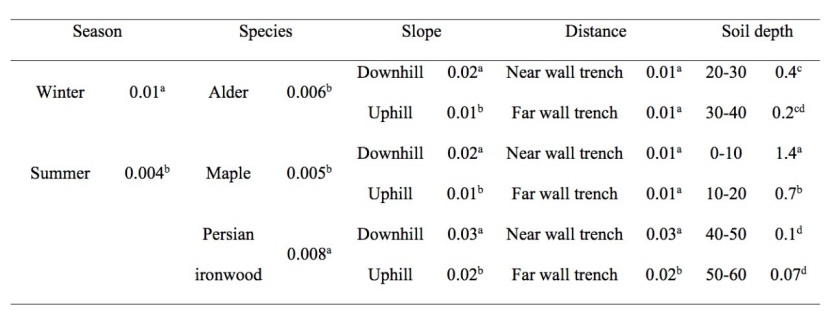
Table 3: Summary of mean comparisons for season, species, slope position, distance from tree stem and soil depth (means with different letters are statistically different (p<0.05)). / Tabelle 3: Zusammenfassung der Mittelwertvergleiche für Jahreszeit, Art, Hanglage, Entfernung vom Baumstamm und Bodentiefe (Mittelwerte mit unterschiedlichen Buchstaben sind statistisch unterschiedlich (p <0.05)).
Root reinforcement decreased in the deeper layers of the soil, i.e. root reinforcement was higher in shallower layers and gradually declined. Additionally, the maximum value of cv was observed in the first 10 cm (Figure 6). Moreover, the findings revealed that root cohesion was significantly higher in winter as compared to the summer corresponding values (p < 0.05).
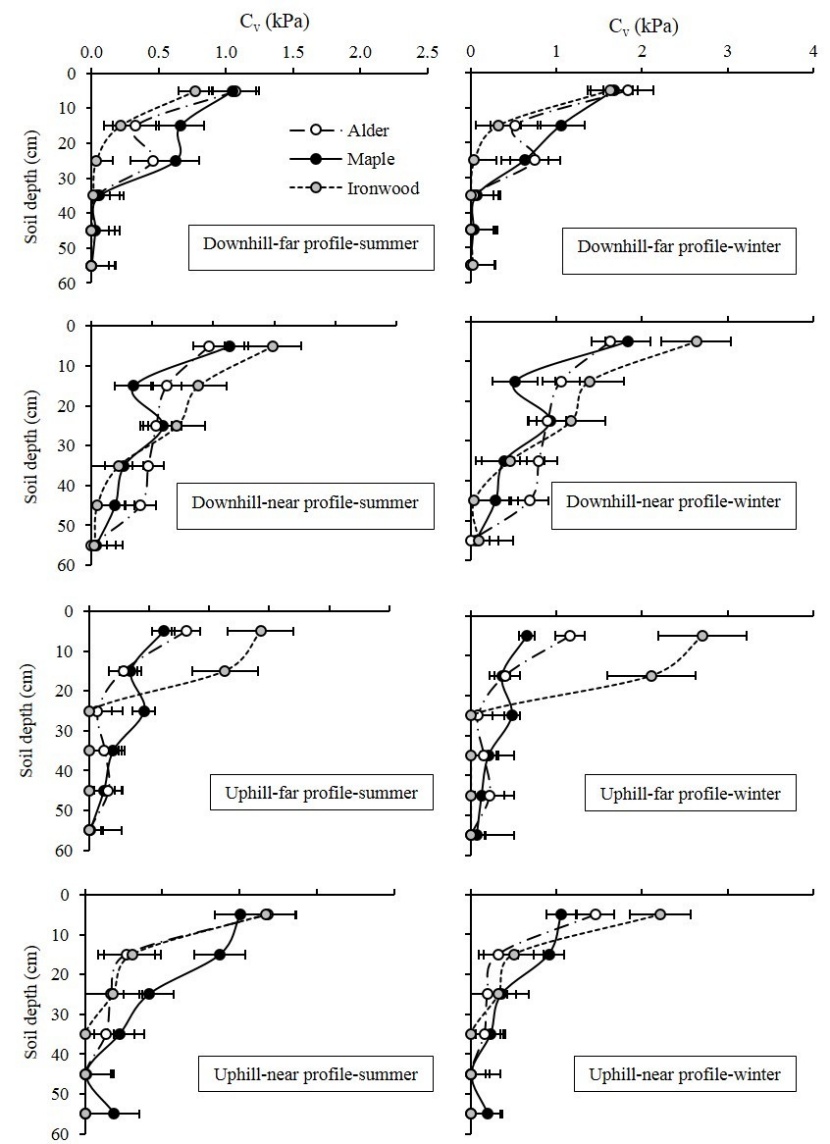
Figure 6: Mean values of root cohesion regarding different treatments (location x season). Different letters means statistically significant differences. / Abbildung 6: Mittelwerte der Wurzelkohäsion in Bezug auf verschiedene Behandlungen (Position x Jahreszeit). Unterschiedliche Buchstaben zeigen statistisch signifikante Unterschiede.
Discussion
The number of roots and RAR values showed high variability among the different species in terms of slope sides, soil depth and distance from the stem. The root density in the first layer of the soil (0-10 cm) was higher than that in the deeper soil layers. The higher density of roots in the topsoil is a normal phenomenon which has been reported by previous researches (Mickovski et al., 2009; Burylo et al., 2011; Adhikari et al., 2013) and has been related to better conditions in the top soil including nutrients, density and aeration (Bischetti et al., 2005; Abdi et al., 2010). Therefore, the highest reinforcement effect of the root system is expected to be in the upper soil layers and this can be prevent shallow landslides. The results indicated that the RAR values declined by increasing the distance from the tree stem, and the RAR value in near profile walls was higher than that in far walls. These findings are consistent with the results of previous studies (e.g., Genet et al., 2008; Ji et al., 2012; Vergani et al., 2014). The reduction factor of the RAR values due to increasing the horizontal distance from the stem was higher than the values reported by Ji et al. (2012). Vergani et al. (2014) discussed that the extent of reduction in the RAR values depended on tree diameter and micro topography and this may justify the different reduction percentages. The amount of the reduction factor can be used efficiently in determining the optimum planting distance of seeds or seedlings in bioengineering projects.
We found large variations in root tensile force and strength on different factors such as season of the year, tree species and root diameter. The relationships between tensile force with strength with root diameter show positive and negative power functions, respectively. Coarse roots need more force to break, although fine roots are stronger in tensile strength (Mao et al., 2012; Vergani et al., 2012). Many researchers reported the same relation (e.g., Genet et al., 2010; Loades et al., 2010; Abdi et al., 2010, 2019) and confirmed the strong dependence of root strength on root size (in our work, the R2 ranges were 0.58 to 0.79). The differences in root chemical composition, especially cellulose content can result in differences in tensile strength (Genet et al., 2005). Moreover, Ye et al. (2017) attributed this relationship to the chemical composition of root tissues and showed that tensile strength was negatively correlated with lignin and the lignin to cellulose ratio, although it was positively correlated with cellulose and holocellulose. However, for tensile force, opposite correlations were reported. The resulting tensile force and strength values in the current study are in the ranges reported in previous studies. The mean tensile force of the three species (42.5-89.4 N) is comparable with the results of Vergani et al. (2016) for some European species (Acer pseudoplatanus: 65 N, Ostrya carpinifolia: 56 N, Fagus sylvatica: 84 N, Fraxinus excelsior: 47 N, and Picea abies: 46 N). However, they were relatively lower than those reported by Chiaradia et al. (2016) for Fagus sylvatica (122.46 N) and Picea abies (70.68 N). The differences between the values presented in the current study and those in the literature may be explained by different responses of plants to different environmental conditions (plasticity) to minimize abiotic and biotic stresses (Boldrin et al., 2017). The mean tensile strength for the three species (22.9-36.0 N) is comparable with that for other hardwood trees in other studies including Quercus robur: 32 MPa and Betula pendula: 38 MPa (Stokes, 2002), Acacia floribunda: 58.1 MPa, Casuarina glauca: 49.6 MPa (Docker and Hubble, 2008), Larrea tridentata: 62.2 MPa, Allenrolfea occidentalis: 35.1 MPa (Adhikari et al., 2013), and Fagus orientalis: 38.2 MPa (Abdi, 2014). It is worth mentioning that tensile strength is considered as an important factor for reinforcement estimation and also for ranking different plant species for bioengineering purposes (Watson and Marden, 2004).
Based on the results, tensile strength in winter was higher than in summer and that is consistent with the results of Norris et al. (2008) and Kazemi et al. (2014) who reported that tensile strength was higher in winter. It was reported that in the leafless period, the root water content decreased. Therefore, roots become more resistant (Coppin and Richards, 1990; Karrenberg et al., 2003). In this regard, Hales and Miniat (2017) found that roots with 50 % less moisture content were more than twice as strong as control roots.
In the Wu model, root reinforcement mainly depends on root tensile strength and RAR or root density at the shear plane (Dumlao et al., 2015). According to the results, root reinforcement decreased with soil depth, and the maximum reinforcement was observed in the upper layer of the soil (0-10 cm). Therefore, the reinforcement effect of vegetation can be more significant for shallow instabilities. Simon and Collison (2002) also reported the maximum root reinforcement in the first 10 cm and with values of 120 kPa in the upper layers, followed by a decrease to 8-12 kPa in deeper layers. In another study, the maximum root reinforcement was reported 60 kPa at the first top layer, and the minimum value of reinforcement was 0.3 kPa at 60 cm depth (Mattia et al., 2005). The results showed that coarse roots have more effect in soil reinforcement due to their higher root area ratio and this is consistent with the results of Schwarz et al. (2013). Regarding the reported values, our results are much lower than those from the literature and the main reason is the low number of roots that intersect the profile walls. This may be a result of dense layers of clay soil in the study area. Moreover, Watson and Marden (2004) reported lower strength values for plantations compared to New Zealand indigenous plant species. As the critical failure plane in the study area was generally in the depth of 1 m, and regarding extremely low values of root reinforcement in the studied stand, it appears that root reinforcement can have an insignificant effect on landslide prevention from the mechanical point of view. Only regarding the cohesion value of soil in the wettest condition in winter (i.e., 1.1 kPa), the reinforcement effect of roots may have some significant effect on landslides prevention.
The data confirmed significant differences between far and near profile walls in the downhill, and this is consistent with other studies showing that root reinforcement decreased by increasing the distance from the tree stem (Schwarz et al., 2010; Vergani et al., 2016) due to the decreasing root density for increasing distances. The effect of distance was not significant in uphill trenches, which may be a result of low density of roots in uphill trenches compared to downhill trenches. The results of this study showed significant differences in reinforcement values among the various species. Persian ironwood compared with the two other plantation species applied more reinforcement in both summer and winter. Watson and Marden (2004) also reported lower strength values for plantations compared to indigenous species. Previous studies in the Hyrcanian forest reported different values of reinforcement for Persian ironwood, i.e. 45 and 0.2 kPa for 0-10 and 60-70 cm soil layers (Abdi et al., 2010), which are much higher than our results. In that study, soil was loamy with lower density compared to the existing study area. This is consistent with the results of previous studies which stated the soil condition would affect reinforcement to a large extent (Genet et al., 2005, 2011; Hales et al., 2013; Zhang et al., 2014; Yang et al., 2016). It is worth to mention that to prevent the risk of failure of individual species, mixtures of species are often suggested in bioengineering projects. It is better that the mixture includes both quick and slow-growing species for providing a plant succession of pioneer species to give immediate protection followed by other species to give a more diversified stand (Morgan and Rickson, 2003).
The findings showed that the root reinforcement values were higher in winter than in summer. Therefore, season of year is one of the parameters which can affect root reinforcement, although there are few studies considering this factor in root reinforcement estimation. The lower water content of roots in the leafless period that has positive effect on resistance may be the reason for the higher reinforcement effect of roots in winter. Form the hydrological point of view, Pollen and Simon (2010) stated that soil cohesion during critical wintertime increased due to soil erodibility; hence, the effects of root reinforcement become more important, especially in areas with a high frequency of rainfall-induced landslides. Considering that most landslides in the study area are rainfall-induced which occur in the leafless period (autumn and winter), hydrological effects of vegetation might not significantly affect soil stability in these seasons with heavy rainfall (Abdi, 2018). Therefore, the mechanical effects of vegetation or root reinforcement can play an important role in soil stabilization. Although the effect of season on reinforcement was significant, the amount of the values was not to the extent that is relevant to prevent landslides (the means were 0.01 kPa in winter versus 0.004 kPa in summer).
After comparing the results with those in other studies, the estimated root cohesion was observed to be much lower than that generally reported in the literature, which is usually > 5 kPa, and calculated using the Wu model (Wu et al., 1979; Simon and Collison, 2002; Mattia et al., 2005; Genet et al., 2008). Field observations showed that the current study site had a thin soil mantle with relatively high soil density (soil bulk density 1.35 g/cm3) that could restrict root system distribution to a great extent. It appears that this is the main reason for the extremely small amount of reinforcement due to the presence of roots. Moreover, we calculated root reinforcement values regarding the empirical correction factor introduced by Preti (2006) in order to correct the overestimation of the Wu model, and this also reduced the reinforcement values. Regarding these low values, it appears that the role of soil reinforcement with tree roots cannot be significant in areas with dense and compacted soil layers which restrict extensive root distribution. Therefore, some researchers suggested measures including site preparation and amelioration to improve soil conditions prior to the establishment phase in bioengineering projects (Morgan and Rickson, 2003).
Concerning the engineering applications, the estimated cohesion supplied due to the presence of roots can be used in analyzing slope stability, assessing the efficiency of erosion control measures, and designing soil bioengineering techniques.
Conclusion
We investigated the root distribution and mechanical properties of three common pioneer tree species of the Hyrcanian forest in Iran. The data collected by this study expands our knowledge on root area ratio (RAR), root tensile strength and root cohesion in alder, maple, and Persian ironwood during two seasons in an area with frequent shallow landslides. RAR values in the downhill sides were higher than those in the uphill sides of trees. RAR of near profile walls of the uphill and downhill sides were significantly higher than those of far walls. We found that RAR decreased by 79 % (downhill) and 59 % (uphill) from 0.5 to 1.0 m distance from the tree stem. Root tensile strength was less in summer than in winter and roots were more resistant in winter, but no significant differences were found among the different species. The relation between tensile strength and root diameter was negative, although it was positive between tensile force and root diameter. Furthermore, Persian ironwood compared with the two other species had more reinforcement effect in both seasons. Moreover, the parameters species, distance from tree stem, soil depth, and season had significant effects on root reinforcement. The presented information can be used to assess the effects of plantations on slope stability and erosion control measures.
Acknowledgements
The authors would like to acknowledge the financial support of the »Iran National Science Foundation (INSF) under the project number 93022486. Also we would like to thank Dr. Jan Friesen (Helmholtz Centre for Environmental Research - UFZ) for editing German parts and the two anonymous reviewers for their detailed comments and suggestions that improved the manuscript.
References
Abdi, E. 2018. Root tensile force and resistance of several tree and shrub species of Hyrcanian forest, Iran. Croatian Journal of Forest Engineering, 39(2): 255–270.
Abdi, E., Azhdary, F., Abdulkhani, A., Sofy Mariv, H. 2014. Tensile strength and cellulose content of Persian ironwood (Parrotia persica) roots as bioengineering material. Journal of Forest Science, 60(1): 425–430.
Abdi, E., Majnounian, B., Genet, M., Rahimi, H. 2010. Quantifying the effects of root reinforcement of Persian Ironwood (Parrotia persica) on slope stability: a case study: hillslope of Hyrcanian forests, Northern Iran. Ecological Engineering, 36: 1409–1416.
Abdi, E., Saleh, HR., Majnounian, B., Deljouei, A. 2019. Soil fixation and erosion control by Haloxylon persicum roots in arid lands, Iran. Journal of Arid Land, 11(1): 86–96.
Adhikari, AR., Gautam, MR., Yu, Z., Imada, S., Achary, K. 2013. Estimation of root cohesion for desert shrub species in the Lower Colorado riparian ecosystem and its potential for streambank stabilization. Ecological Engineering, 51: 33–44.
Bassanelli, C., Bischetti, GB., Chiaradia, EA., Rossi, L., Vergani, C. 2013. The contribution of chestnut coppice forests on slope stability in abandoned territory: A case study. Journal of Agricultural Engineering, 44: 68–73.
Bischetti, GB., Chiaradia, EA., D’agostino, V., Simonato, T. 2010. Quantifying the effect of brush layering on slope stability. Ecological Engineering, 36(3): 258–264.
Bischetti, GB., Chiaradia, EA., Simonato, T., Speziali, B., Vitali, B., Vullo, P., Zocco, A. 2005. Root strength and root area ratio of forest species in Lombardy (North¬ern Italy). Plant and Soil, 278: 11–22.
Boldrin, D., Leung, AK., Bengough, AG. 2017. Root biomechanical properties during establishment of woody perennials. Ecological Engineering, 109: 196–206.
Bordoloi. S., Yamsani, SK., Garg, A., Sreedeep, S., Borah, S. 2015. Study on the efficacy of harmful weed species Eicchornia crassipes for soil reinforcement. Ecological Engineering, 85: 218–222.
Burylo, M., Hudek, C., Rey, F. 2011. Soil reinforcement by the roots of six dominant species on eroded mountainous marly slopes (Southern Alps, France). Catena, 84: 70–78.
Chiaradia, EA., Bischetti, GB., Vergani, C. 2012. Incorporating the effect of root systems of forest species into spatially distributed models of shallow landslides. International Journal of Forest, Soil, and Erosion, 2(3): 107–118.
Chiaradia, EA., Vergani, C., Bischetti, GB. 2016. Evaluation of the effects of three European forest types on slope stability by field and probabilistic analyses and their implications for forest management. Forest Ecology and Management, 370: 114–129.
Chiatante, D., Sarnataro, M., Fusco, S., Di Dorio, A., Scippa, GS. 2003. Modification of root morphological parameters and root architecture in seedlings of Fraxinus ornus L. and Spartium junceum L. growing on slopes. Plant Biosystems, 137: 47–56.
Collins, AJ. 2001. The role of willow root architecture and character in root reinforcement potential, doctoral thesis, University of Nottingham, Nottingham, U.K.
Coppin, NJ., Richards, IG. 1990. Use of vegetation in Civil Engineering. Butterworth, London.
De Baets, S., Poesen, J., Reubens, B., Wemans, K., De Baerdemaeker, J., Muys, B. 2008. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant and Soil, 305(1–2): 207–226.
Deljouei, A., Sadeghi, SMM., Abdi, E., Bernhardt-Romermann, M., Pascoe, EL., Marcantonio, M. 2018. The impact of road disturbance on vegetation and soil properties in a beech stand. Hyrcanian forest. European Journal of Forest Research, 137(6): 759–770.
Docker, BB., Hubble, TCT. 2008. Quantifying root-reinforcement of river bank soils by four Australian tree species. Geomorphology, 100: 401–418.
Dumlao, MR., Ramananarivo, S., Goyal, V., De Jong, JT., Waller, J., Silk, WK. 2015. The role of root development of Avena fatua in conferring soil strength. American Journal of Botany, 102(7): 1050–1060.
Genet, M., Kokutse, N., Stokes, A., Fourcaud, T., Cai, X., Ji, J., Mickovski, SB. 2008. Root reinforcement in plantations of Cryptomeria japonica D. Don: effect of tree age and stand structure on slope stability. Forest Ecology and Management, 256: 1517–1526.
Genet, M., Li, M., Luo, T., Fourcaud, T., Clément-Vidal, A., Stokes, A. 2011. Linking carbon supply to root cell-wall chemistry and mechanics at high altitudes in Abies georgei. Annals of Botany, 107: 311–320.
Genet, M., Stokes, A., Fourcaud, T., Hu, X., Lu, Y. 2006. Soil fixation by tree roots: changes in root reinforcement parameters with age in Cryptomeria japonica D. Don. plantations. In: Disaster Mitigation of Debris Flows, Slope Failures and Landslides. pp. 535–542. Universal Academy Press, Inc., Tokyo, Japan.
Genet, M., Stokes, A., Fourcaud, T., Norris, JE. 2010. The influence of plant diversity on slope stability in moist evergreen deciduous forest. Ecological Engineering, 36: 265–275.
Genet, M., Stokes, A., Salin, F. 2005. The influence of cellulose content on tensile strength in tree roots. Plant and Soil, 278: 1–9.
Gentile, F., Elia, G., Elia, R. 2010. Analysis of the stability of slopes reinforced by roots. In: Brebbia, CA., Carpi, A. (Eds.) Design and Nature V: Comparing Design in Nature with Science and Engineering. WIT Transactions Ecology Environment, 138: 189–200.
Giadrossich, F., Cohen, D., Schwarz, M., Seddaiu, G., Contran, N., Lubino, M., Valdés-Rodrŕguez, OA., Niedda, M. 2016. Modeling bio-engineering traits of Jatropha curcas L., Ecological Engineering, 89: 40–48.
Gilardelli, F., Vergani, C., Gentili, R., Bonis, A., Pierre, C., Sandra, C., Chiaradia, EA. 2017. Root Characteristics of herbaceous species for topsoil stabilization restoration in projects. Land Degradation and Development, 28: 2074–2085.
Gray, DH., Sotir, RB. 1996. Biotechnical and Soil Bioengineering Slope Stabilization. A Practical Guide for Erosion Control. John Wiley & Sons.
Hales, TC., Cole-Hawthorne, C., Lovell, L., Evans, S. 2013. Assessing the accuracy of simple field based root strength measurements. Plant and Soil, 372: 553–565.
Hales, TC., Ford, CR., Hwang, T., Vose, JM., Band, LE. 2009. Topographic and ecologic controls on root reinforcement. Journal of Geophysical Research, 114: DOI: 10.1029/2008JF001168.
Hales, TC., Miniat, CF. 2017. Soil moisture causes dynamic adjustments to root reinforcement that reduce slope stability. Earth Surface Processes and Landforms, 42(5): 803–813.
Heidarian, T., Ghasemi Aghbahs, F., Pazhouhan, I. 2018. The role of native species of Quercus brantti and xin soil reinforcement in Zagros forests. Austrian Journal of Forest Science, 4: 283–296
Hytiris, N., Fraser, M., Mickovski, SB. 2015. Enhancing slope stability with vegetation. International Journal of GEOMATE, 9(18): 1477–1482.
Ji, J., Kokutse, N., Genet, M., Fourcaud, T., Zhang, Z. 2012. Effect of spatial variation of tree root characteristics on slope stability. A case study on black locust (Robinia pseudoacacia) and arborvitae (Platycladus orientalis) stands on the Loess Plateau, China. Catena, 92: 139–154.
Karrenberg, S., Blaser, S., Kollmann, J., Speck, T., Edwards, PJ. 2003. Root anchorage of saplings and cuttings of woody pioneer species in a riparian environment. Functional Ecology, 17: 170–177.
Kazemi, M., Abdi, E., Majnounian, B., Yousef Zadeh, H. 2014. The effect of season on resistance of Persian oak (Quercus persica) roots (Case study: Tabarok, Bazaft basin). Iranian Journal of Forest, 6(4): 435–444.
Konopka, B. 2017. Foliage and fine root litter: A comparative study in young, natural regenerated stands of European beech and Norway spruce. Austrian Journal of Forest Science, 134(2): 99–118.
Loades, KW., Bengough, AG., Bransby, MF., Hallett, PD. 2010. Planting density influence on fibrous root reinforcement of soils. Ecological Engineering, 36: 276–284.
Mao, Z., Saint-André, L., Genet, M., Mine, FX., Jourdan, C., Rey, H., Courbaud, B., Stokes, A. 2012. Engineering ecological protection against landslides in diverse mountain forests: choosing cohesion models. Ecological Engineering, 45: 55–69.
Mattia, C., Bischetti, GB., Gentile, F. 2005. Biotechnical characteristics of root systems of typical Mediterranean species. Plant and Soil, 278: 23–32.
Mickovski, SB., Hallett, PD., Bransby, MF., Davies, MCR., Sonnenberg, R., Bengough, AG. 2009. Mechanical reinforcement of soil by willow roots: impacts of root properties and root failure mechanism. Soil Science Society of America Journal, 73: 1276–1285.
Morgan, RP., Rickson, RJ. 2003. Slope stabilization and erosion control: a bioengineering approach. Taylor & Francis.
Naghdi, R., Maleki, S., Abdi, E., Mousavi, R., Nikooy, M. 2013. Assessing the effect of Alnus roots on hillslope stability in order to use in soil bioengineering. Journal of Forest Science, 59(11): 417–423.
Nilaweera, NS. 1994. Effects of Tree Roots on Slope Stability: The Case of Khao Luang Mountain Area, So Thailand. Doctoral Thesis. Asian Institute of Technology, Bangkok.
Norris, JE., Stokes, A., Mickovski, SB., Cammeraat, E., Van Beek, R., Nicoll, BC., Achim, A. 2008. Slope Stability and Erosion Control: Ecotechnological Solutions, 283 p.
Nyambane, OS., Mwea, SK. 2011. Root tensile strength of three typical plant species and their contribution to soil shear strength; a case study, Sasumua Backslope, Nyandarua District, Kenya. Journal of Civil Engineering Research and Practice, 8(1): 57–73.
Operstein, V., Frydman, S. 2000. The influence of vegetation on soil strength. Ground Improvement, 4: 81–89.
Pollen, N., Simon, A. 2005. Estimating the mechanical effects of riparian vegetation on streambank stability using a fiber bundle model. Water Resources Research, DOI: 10.1029/2004WR003801.
Pollen, N., Simon, A. 2010. Hydrologic and hydraulic effects of riparian root networks on streambank stability: Is mechanical root-reinforcement the whole story? Geomorphology, 116: 353–362.
Preti, F. 2006. Stabilit`a dei versanti vegetati, Cap. 10, in: Manuale 3 d’Ingegneria Naturalistica Sistemazione dei versanti, edited by: Sauli, G., Cornelini, P., Preti, F., Regione Lazio, Roma, pp. 137–168.
Preti, F. 2013. Forest protection and protection forest: tree root degradation over hydrological shallow landslides triggering. Ecological Engineering, 61: 633–645.
Reubens, B., Poesen, J., Danjon, F., Geudens, G., Muys, B. 2007. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: a review. Trees, 21: 385–402.
Rodriguez, OA., Niedda, M. 2016. Modeling bio-engineering traits of Jatropha curcas L. Ecological Engineering, 89: 40–48.
Roering, JJ., Schmidt, KM., Stock, JD., Dietrich, WE., Montgomery, DR. 2003. Shallow landsliding, root reinforcement, and the spatial distribution of trees in the Oregon Coast Range. Canadian Geotechnical Journal, 40: 237–253.
Saxena, D., Flores, S., Stotzky, G. 1999. Insecticidal toxin in root exudates from Bt corn. Nature, 402: 480.
Schiechtl, HM. 1980. Bioengineering for Land Reclamation and Conservation. University of Alberta Press, Edmonton, Alberta, Canada.
Schmaltz, EM., Steger, S., Glade, T. 2017. The influence of forest cover on landslide occurrence explored with spatio-temporal information. Geomorphology, 290: 250–264.
Schmidt, KM., Roering, JJ., Stock, JD., Dietrich, WE., Montgomery, DR., Schaub, T. 2001. The variability of root cohesion as an influence on shallow landslide susceptibility in the Oregon Coast Range. Canadian Journal of Forest Research, 38: 995–1024.
Schwarz, M., Cohen, D., Or, D. 2012. Spatial characterization of root reinforcement at stand scale: Theory and case study. Geomorphology, 171–172: 190–200.
Schwarz, M., Giadrossich, F., Cohen, D. 2013. Modeling root reinforcement using a root-failure Weibull survival function. Hydrology and Earth System Science, 17(11): 4367–4377.
Schwarz, M., Lehmann, P., Or, D. 2010. Quantifying lateral root reinforcement in steep slopes from a bundle of roots to tree stands. Earth Surface Processes and Landforms, 35: 354–367.
Simon, A., Collison, AJC. 2002. Quantifying the mechanical and hydrologic effects of riparian vegetation on streambank stability. Earth Surface Processes and Landforms, 27: 527– 546.
Stokes, A. 2002. The biomechanics of tree root anchorage. In: Waisel Y, Eshel A, Kafkaki U (Ed.) “Plant Roots- The Hidden Half”. Plenum Publishing, New York, pp. 175–186.
Stokes, A., Sotir, R, Chen, W., Chestem, M. 2010. Soil bio- and ecoengineering in China: Past experience and future priorities. Ecological Engineering, 36: 247–257.
Tardio, G., González-Ollauri, A., Mickovski, SB. 2016. A noninvasive preferential root distribution analysis methodology from a slope stability approach. Ecological Engineering, 97: 46–57.
Tardio, G., Mickovski, SB. 2015. Method for synchronisation of soil and root behaviour for assessment of stability of vegetated slopes. Ecological Engineering, 82: 222–230.
Tosi, M. 2007. Root tensile strength relationships and their slope stability implications of three shrub species in the northern Apennines (Italy). Geomorphology, 87(4): 268–283.
Vergani, C., Chiaradia, E., Bischetti, G. 2012. Variabil¬ity in the tensile resistance of roots in Alpine forest tree species. Ecological Engineering, 46: 43–56.
Vergani, C., Giadrossich, F., Buckley, P., Conedera, M., Pividori, M., Salbitano, F., Rauch, HS., Lovreglio, R., Schwarz, M. 2017. Root reinforcement dynamics of European coppice woodlands and their effect on shallow landslides: A review. Earth Science Review, 167: 88–102.
Vergani, C., Schwarz, M., Cohen, D., Thormann, JJ., Bischetti, GB. 2014. Effects of root tensile force and diameter distribution variability on root reinforcement in the Swiss and Italian Alps. Canadian Journal of Forest Research, 44: 1426–1440.
Vergani, C., Schwarz, M., Soldati, M., Corda, A., Giadrossich, F., Chiaradia, EA., Morando, P., Bassanelli, C. 2016. Root reinforcement dynamics in subalpine spruce forests following timber harvest: a case study in Canton Schwyz, Switzerland. Catena, 143: 275–288.
Waldron, LJ. 1977. The shear stress resistance of root-permeated homogeneous and stratified soil. Soil Science Society of America Proceeding, 41: 843–849.
Watson, AJ., Mardern, M. 2004. Live root-wood tensile strengths of some common New Zealand indigenous and plantation tree species. New Zealand Journal of Forestry Science, 34(3): 344–353.
Wu, TH., Mckinnell, WO., Swanston, DN. 1979. Strength of tree roots and landslides on Prince-Of-Wales-Island, Alaska. Canadian Geotechnical Journal, 16(1): 19–33.
Wu, W. 2016. Recent Advances in Modelling Landslides and Debris Flows. Springer, Netherlands, 322 p.
Yang, Y., Chen, L., Li, N., Zhang, Q. 2016. Effect of root moisture content and diameter on root tensile properties. Plos One, 11: e0151791. DOI:10.1371/journal.pone.0151791.
Ye, C., Guo, Z., Li, Z., Cai, C. 2017. The effect of Bahiagrass roots on soil erosion resistance of Aquults in subtropical China. Geomorphology, 285: 82–93.
Zare, M., Pourghasemi, HR., Vafakhah, M., Pradhan, B. 2013. Landslide susceptibility mapping at Vaz Watershed (Iran) using an artificial neural network model: a comparison between multilayer perceptron (MLP) and radial basic function (RBF) algorithms. Arab Journal of Geosciences, 6(8): 2873–2888.
Zhang, C., Chen, L., Jiang, J. 2014. Why fine tree roots are stronger than thicker roots: the role of cellulose and lignin in relation to slope stability. Geomorphology, 206: 196–202.



