Schlüsselbegriffe: Allelopathie, Waldboden, Saisonalität, Eucalyptus grandis x E. urophylla, Erythrophleum fordii, Streuzersetzung, Aufforstung, nicht-heimische Baumarten
Abstract
Accumulation of phenolic acids in monocropping plantations led to declining soil fertility and reduced productivity due to allelopathic effects. The effects of mixed versus mono-specific plantations on the seasonal turnover of phenolic acids in soil remains unclear. This study explored in plantations of Eucalyptus grandis × Eucalyptus urophylla and Erythrophleum fordii Oliv. (mixed) versus E. grandis × E. urophylla only (pure) the effect of species mixture on the phenolic acid content in the soil. We analyzed both total phenolic acid content as well as six phenolic acids separately and our measurements were conducted over four seasons and in an entire year. We expected that the mixed plantation would exhibit lower accumulation of both total and individual phenolic acids in soil versus the pure plantation. Contrary to our hypothesis, the total content of phenolic acids in the mixed plantation was significantly (p < 0.001) higher than that in the pure Eucalyptus plantation in all seasons. We also observed a significant effect of stand type (mixed or pure), season, and interaction between stand type and season on total soil phenolic acid content (p < 0.001). This may cause an enhanced allelopathic effect of phenolic acids in the soil of the mixed stand. In terms of the individual phenolic acids, the mixed plantation resulted in significantly higher (p < 0.001) p-hydroxybenzoic acid and vanillic acid contents in all seasons, as compared with the pure Eucalyptus plantation. Species mixture appears to reduce the content of ferulic acid, coumaric acid, benzoic acid, and salicylic acid in most seasons. A significant effect (p < 0.001) of stand type, season, and their interaction on the content of all individual phenolic acids in the soil, with the exception of benzoic acid, was observed. This study thus strongly suggests that both total and individual phenolic acids should be studied to evaluate the possible allelopathic effects in forest ecosystems and verify that species mixture can also increase phenolic acid concentration.
Zusammenfassung
Die Akkumulation von Phenolsäure in Monokulturen wurde wegen ihres allelopathischen Effekts in Zusammenhang mit Verminderung von Bodenfruchtbarkeit und -produktivität gebracht. Die Auswirkungen von Baumartenmischung auf die jahreszeitlichen Schwankungen des Phenolsäure-Gehalts im Boden ist wenig erforscht. Diese Studie untersucht den Effekt von Baumartenmischung auf den Phenolsäure-Gehalt im Boden von gemischten Eucalyptus grandis × Eucalyptus urophylla und Erythrophleum fordii Oliv. Plantagen im Vergleich zu einem reinen E. grandis × E. urophylla Bestand. Wir untersuchten sowohl den gesamten Phenolsäure-Gehalt als auch den Gehalt von sechs einzelnen Phenolsäuren während vier Jahreszeiten über ein ganzes Jahr. Wir haben erwartet, dass sowohl der totale als auch der individuelle Phenolsäure-Gehalt im Reinbestand höher wäre als im Mischbestand. Entgegen unserer Hypothese war der totale Phenolsäure-Gehalt in der gemischten Bepflanzung signifikant (p < 0.001) höher als der Phenolsäure-Gehalt in dem Eukalyptus Reinbestand in allen Jahreszeiten. Es zeigte sich auch eine signifikanter Effekt von Bepflanzungstyp (gemischt oder rein), Jahreszeit und Interaktion zwischen Bepflanzungstyp und Jahreszeit auf den totalen Phenolsäure-Gehalt (p < 0.001). Das kann einen verstärkten allelopathischen Effekt der Phenolsäure im Boden des Mischbestandes zur Folge haben. Bei den einzelnen Phenolsäuren hat der Mischbestand einen signifikant höheren Gehalt (p < 0.001) von p-Hydroxybenzoesäure und Vanillinsäuregehältern in allen Jahreszeiten als der Reinbestand. Gemischte Bepflanzung reduzierte den Gehalt von Ferulsäure, Coumarinsäure, Benzoesäure und Salicylsäure in den meisten Jahreszeiten. Eine signifikante Wirkung (p < 0.001) von Bepflanzungstyp, Jahreszeit und deren Interaktion auf den Gehalt der einzelnen Phenolsäuren (ausgenommen Benzoesäure) wurde beobachtet. Unsere Studie legt nahe, dass sowohl die totalen als auch einzelnen Phenolsäure-Gehälter im Boden erforscht werden müssen, um mögliche allelopathische Effekte von Phenolsäuren in Wäldern zu verstehen und um zu verifizieren, ob Baumartenmischung auch zu einer Erhöhung des Phenolsäure-Gehalts führen kann.
1. Introduction
Eucalyptus spp. has been widely cultivated in many provinces of southern China, because of its fast growth, good adaptability, and high economic efficiency and in turn has become an important component of the forest economy. According to the results of the Ninth National Forest Resource Inventory Report, the area of Eucalyptus plantations in China has reached 4.45 million hectares (The State Forestry Administration of the People’s Republic of China 2014). In southern China, 60 % – 70 % of Eucalyptus plantations are short-rotation plantations, of which 50 % – 60 % use monocropping (Wen 2005).
With the increasing share of monocropping in Eucalyptus plantations, concerns about the potential environmental impacts become more frequent. Short rotations may lead to loss of soil fertility and productivity and affect biodiversity negatively (Jagger and Pender 2000, 2003; Laclau et al. 2010). Allelopathic effects inhibiting the growth of plants and microorganisms in forest ecosystems can be caused by certain chemical substances produced by leaching, litter decomposition, and/or root exudation and can lead to further loss of biodiversity (Bolte et al. 1984; Zeng and Li 1997). The leachate from Eucalyptus leaves can reduce the growth of Eucalyptus viminalis seedlings by 80 % – 85 % and the leaching effect can lead to the formation of a circular inhibition zone under the canopy (Zhai and Jia 1993). Studies have also shown that phenolic acids are important allelopathic substances in the soils of Eucalyptus grandis plantations (Chen et al. 2002; Wang et al. 2006; Wang et al. 2007).
Phenolic acids are secondary metabolites of plants. Most phenolic acids have strong allelopathic effects and can affect the growth and development of plants of the same but also other species through soil regulatory effects (Blum and Dalton 1985). Several studies have shown that high concentration of phenolic acids can severely inhibit plant growth (Iannucci et al. 2013; Patrick 1971; Patterson 1981; Zhu et al. 2003). Forest and agricultural soils contain various phenolic acids as well as other toxic and non-toxic organic compounds that are capable of interacting with soil organisms and roots (Blum 1996; Inderjit 1996). Measurements of soil ester-linked phenolic acids have revealed a large range of phenolic compounds including p-hydroxybenzoic acid, vanillic acid, syringic acid, coumaric acid, and ferulic acid (Martens 2002). Understanding the ecological impact of the various phenolic acids and their interaction effects is essential for understanding the phenomenon of allelopathy (Inderjit 1996). For instance, the inhibitory effect of ferulic acid on leaf expansion, transpiration, water use efficiency, and leaf area has been studied for cucumber seedlings (Blum and Dalton 1985; Blum and Gerig 2005). The inhibitory effect of cinnamic acid derivatives (ferulic acid and p-coumaric acid) was two- to five-fold higher than that of benzoic acid derivatives (p-hydroxybenzoic acid and vanillic acid) (Blum and Gerig 2005).
The transformation of pure into mixed forests might solve the ecological problems associated with pure forests like increase of pests and diseases, decreasing biodiversity, and declining soil fertility (Bravo-Oviedo et al. 2014; Carnol et al. 2014; Knoke et al. 2008; Xavier et al. 2011). Since 2006, the establishment of mixed plantations of Eucalyptus with other tree species has become more common in southern China, as ecological problems of pure Eucalyptus plantations get more attention. Erythrophleum fordii Oliv. is a valuable native hardwood species that is also nitrogen-fixing (Yao et al. 2014) and is now frequently used for mixed planting with Eucalyptus. Previous studies mostly focused on tree species mixing in terms of tree growth (Bouillet et al. 2013; Mason and Connolly 2014), light-use efficiency (Forrester and Albrecht 2014), water use efficiency (Forrester et al. 2010), biodiversity (Chauvat et al. 2011), and soil nutrient changes (Bouillet et al. 2008; Forrester et al. 2005). Few studies have focused on tree species mixing in terms of phenolic acid allelopathy in soil. To our knowledge, there has been no research on soil phenolic acids in mixed plantations of Eucalyptus and Erythrophleum fordii. The allelopathic effect of phenolic acid is closely related to the concentration and type of phenolic acid. In this study, we studied the content of six phenolic acids (p-hydroxybenzoic acid, vanillic acid, ferulic acid, benzoic acid, coumaric acid, and salicylic acid) over four seasons were studied in the soils of a pure Eucalyptus plantation and a mixed plantation of Eucalyptus with Erythrophleum fordii. We like to investigate the effect of mixed planting of Eucalyptus and Erythrophleum fordii on the turnover of phenolic acids in forest soil and provide data for evaluating the allelopathy of phenolic acids based on their content in the soil of the two types of plantation in the future. Accumulation of phenolic acid might be one of the important triggers of soil fertility decline (Tan et al. 2008; Ye et al. 2006; Zhou et al. 2012). Since mixed forests often better maintain soil fertility than pure forests, we expected that the mixed plantation would have lower accumulation of both total and individual phenolic acids in soil versus the pure plantation. To test this hypothesis, fixed sampling points were set in the two plantations and soil samples were collected in March, June, September, and December 2014. The content of each type of phenolic acid was then determined by high-performance liquid chromatography (HPLC).
2. Materials and methods
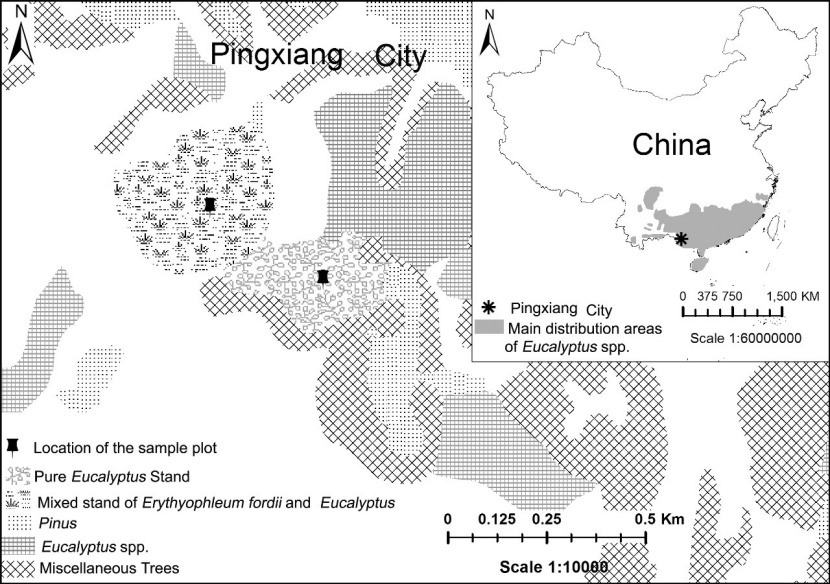
Figure 1: The location of the experiment site and the forests surrounding the experiment site. / Abbildung 1: Die Lage des Versuchsflächen und die die nähere Umgebung.
2.1. Study area and site description
The experimental site was located at the Experimental Center of Tropical Forestry of the Chinese Academy of Forestry (22°7’N, 106°42’E). The Center is located in Pingxiang, Guangxi Province, China and has a south subtropical monsoon climate with an average annual rainfall of 1500 mm and an annual average temperature of 21 °C. A pure Eucalyptus grandis × E. urophylla plantation (denoted as “pure”) and a mixed plantation of E. grandis × E. urophylla and Erythrophleum fordii (denoted as “mixed”) with the same environmental factors were selected for experiments (Figure 1).
There was a 2:1 ratio between E. grandis × E. urophylla and Erythrophleum fordii in the mixed plantation. The planting patterns of the pure and mixed plantations are shown in Figure 2.
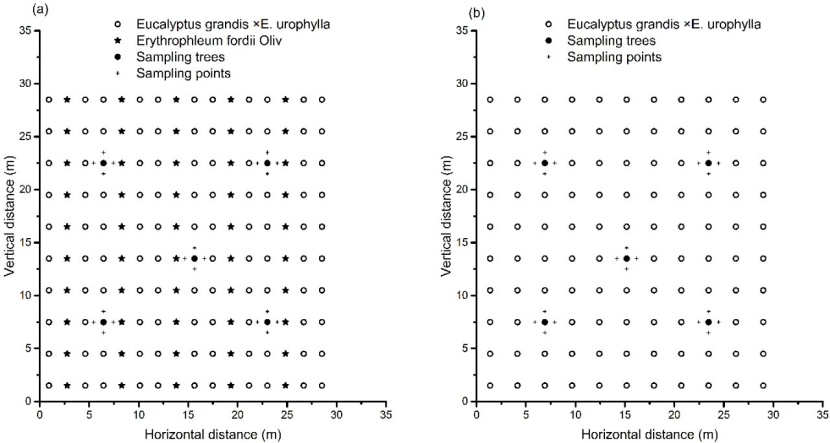
Figure 2: Layout of trees in (a) the mixed (Eucalyptus grandis × E. urophylla and Erythrophleum fordii Oliv.) and (b) the pure (E. grandis × E. urophylla) plantation stands. / Abbildung 2: Übersichtsplan der Standorte der Bäume in den (a) gemischten (Eucalyptus grandis × E. urophylla und Erythrophleum fordii Oliv.) und (b) reinen (E. grandis × E. urophylla) Bepflanzungen.
The pure and mixed stands were adjacent to one another and establishment with seedlings in April 2009 (stand age in 2014 was 5 years). Prior to 2009 this area was planted with Pinus massoniana. The soil type of both plantations was classified as red soil in the Chinese soil classification, equivalent to Oxisol in the USDA Soil Taxonomy (Wang et al. 2013). The slope aspect of the two plantations was northwest, and the slope position of the two plantations was 20-25˚. The main understory species of the two plantations were Microstegium vagans, Miscanthus floridulus, and Thysanolaena maxima. The basic properties of the two plantations are summarized in Table 1.
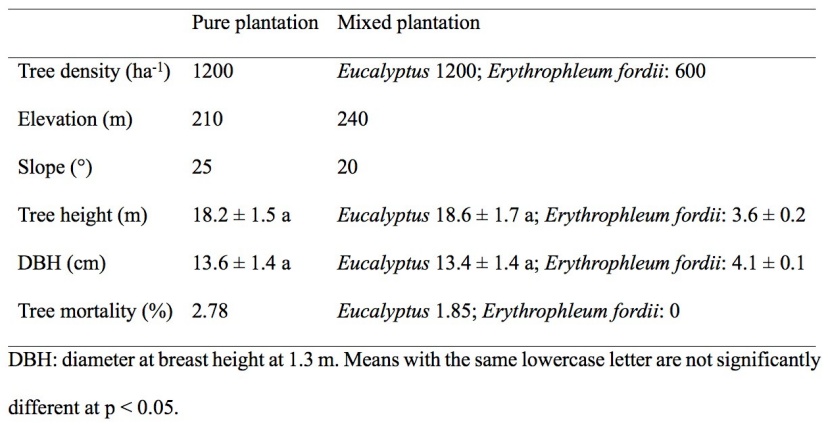
Table 1: Basic properties on the studied mixed (Eucalyptus grandis × E. urophylla and Erythrophleum fordii Oliv.) and pure (E. grandis × E. urophylla) plantations in year 2014. / Tabelle 1: Grundlegende Informationen über die untersuchten gemischten (Eucalyptus grandis × Eucalyptus urophylla und Erythrophleum fordii Oliv.) und reinen (E. grandis × E. urophylla) Plantagen im Jahr 2014.
2.2. Sample collection
In March 2014, a 900 m2 (30 m × 30 m) plot was marked in the center of the selected pure and mixed plantations. The diameter at breast height (DBH) of every E. grandis × E. urophylla and Erythrophleum fordii tree in the plot was measured, and the average value and tree mortality was calculated for each plot by comparing to the initial tree density (Table 1). Five E. grandis × E. urophylla trees with an average DBH were selected from each plot, and then four fixed sampling points were set at a distance of 1.0 m away from the base of the trunk of each selected tree. The fixed sampling points were labelled, and soil samples were collected down to a depth of 30 cm using a soil auger with an inner diameter of 10 cm. The distributions of sample trees and sampling points are shown in Figure 2. In order to retain the phenolic acids in the soil, all soil samples were quickly placed in sterile glass containers and stored in a portable cooler. In the laboratory, the soil samples for each stand were mixed and stored in a refrigerator for later use. In June, September, and December 2014, soil samples were collected from the fixed sampling points of the two stands in the same way.
2.3. Analyzing phenolic acid content
Within two days of sampling in each season, the phenolic acid content was analysed. To avoid the degradation of phenolic acid by microorganisms in the soil, the soil samples were sterilized by cobalt-60 irradiation to a total dose of 30 kGy (Wolf et al. 1989). Then, 25.00 g of sterilized pure plantation soil and mixed plantation soil were weighed into different sterilized centrifuge tubes, 25 mL of 1 mol/L NaOH was added, and the mixture was allowed to stand for 24 h. The mixture was then shaken for 30 min and centrifuged at 4000 r/min; the centrifugate was filtered and acidified with 12 M hydrochloric acid to a pH of 2.5. After 2 h, the sample was centrifuged to remove humic acid and was then analyzed by HPLC.
Three replicate experiments were performed for each soil sample to determine the content of six phenolic acids (p-hydroxybenzoic acid, vanillic acid, ferulic acid, benzoic acid, coumaric acid, and salicylic acid) in all soil samples. HPLC analysis was conducted for the phenolic acid standards, and the peak area of the phenolic acids was plotted against the concentrations of the standard. The linear equation of the phenolic acid concentration and the peak area was obtained by linear regression. The phenolic acids in the solution to be determined and their concentrations were identified and calculated by comparing the peak retention times and peak areas of the phenolic acid standard solutions, respectively (Dalton et al. 1989). Phenolic acid standards were prepared using ultra-pure water, and all solutions for chromatographic analysis were centrifuged at room temperature for 10 min at 4000 r/min, followed by filtration through a 0.22 μm membrane (Wang 2010). Chromatographic analysis was carried out using an HPLC system with a Waters 2487 HPLC Absorbance UV-Vis Detector and a μBondapak C18 3.9 × 300 mm column (Waters Corp., Milford, MA, USA). The UV detector was used with a detection wavelength of 280 nm, and the column temperature was set at 30 °C. The mobile phase was acetonitrile and ultrapure water (10 % acetonitrile and 1 % glacial acetic acid). The injection volume was 20 μL, and the flow rate was 1.1 mL/min.
2.4. Statistical analysis
Basic statistical data analysis was performed using Microsoft Office Excel 2010 (Microsoft Corp., Redmond, WA, USA). Figure 1 was created with ArcGIS 10.0 (Environmental Systems Research Institute Inc., Redlands, CA, USA), Figures 2–4 were created with the Origin 9.1 software (OriginLab Corp., Northampton, MA, USA), and analysis of variance (ANOVA) was performed using the SAS systems for windows Version 8.0 (SAS Institute Inc., Cary, NC, USA).
3. Results and discussion
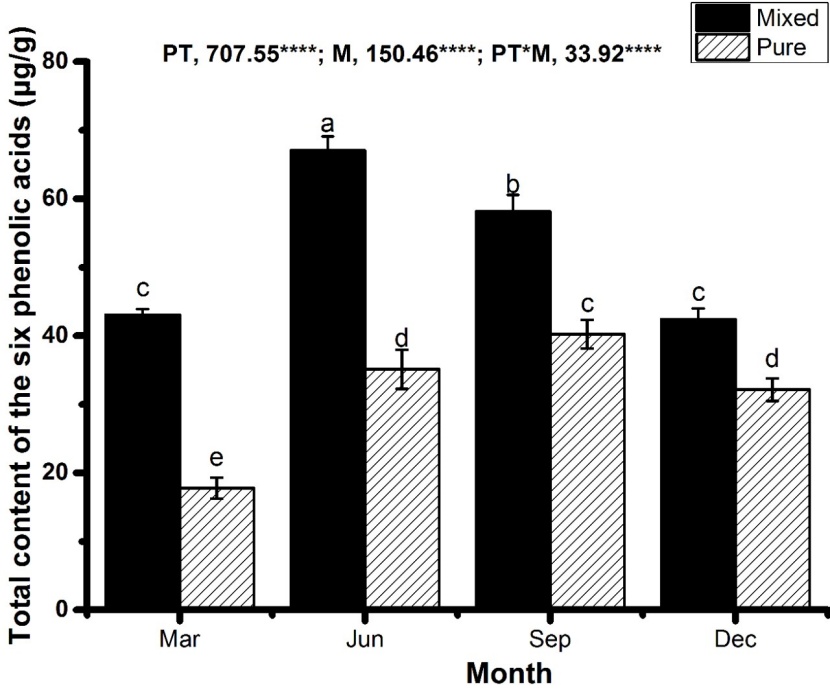
Figure 3: The total contents of six phenolic acids in the soil of the mixed (Eucalyptus grandis × E. urophylla and Erythrophleum fordii Oliv.) and pure (E. grandis × E. urophylla) plantations in different seasons (March, June, September, and December). Data are presented as mean ± SEM. Bars with different lowercase letters indicate significant difference at p < 0.05. F values from the two-way ANOVA analysis are given above each chart. PT, plantation type; M, month; * p < 0.05, *** p < 0.001. / Abbildung 3: Der Gesamtgehalt der sechs Phenolsäuren in den gemischten (Eucalyptus grandis × E. urophylla und Erythrophleum fordii Oliv.) beziehungsweise reinen (E. grandis × E. urophylla) Bepflanzungen in den verschiedenen Jahreszeiten (März, Juni, September, und Dezember). Daten sind dargestellt als Mittelwert ± SEM. Mit unterschiedlichen Kleinbuchstaben bezeichnete Säulen zeigen signifikante Unterschiede bei p < 0.05. Oberhalb eines jeden Diagrammes sind die F-Werte der zweifachen ANOVA angegeben. PT: Bepflanzungstyp; M: Monat: * p < 0.05, ***: p < 0.001.
3.1. Total phenolic acid content in soil
The total content of phenolic acids in the mixed plantation soil was significantly higher than that in the pure plantation soil in every season (Figure 3).
A significant (p < 0.001) effect of stand-type, season, and the interaction between stand type and season was observed on the total phenolic acid content in the soil (Figure 3). The increase in total phenolic acid might be attributed to five reasons. First, microorganisms in the mixed plantation soil might be more abundant than those in the pure plantation (Behera and Sahani 2003; Wang and Wang 2008), which promote the decomposition of litter in the stand and produce more phenolic acid (Kuiters 1990; Kuiters and Sarink 1986). Second, the secretion from Erythrophleum fordii roots is a new source of phenolic acid in the mixed plantation soil, compared with the pure plantation soil. Third, greater stand density in the mixed plantation can lead to more litter input and/or change the litter decomposition (Fang et al. 2015; Liu et al. 2006; Meng et al. 2011; Xing et al. 2019). Fourth, the carbon:nitrogen ratio of the litter might change due to addition of the Erythrophleum fordii litter with higher N concentration, which may promote litter decomposition in the mixed plantation versus that in the pure plantation (Wang et al. 2016). Fifth, the nitrogen fixation of the rhizobia of Erythrophleum fordii roots can enhance the activity of soil microorganisms in the mixed plantation, which promotes the decomposition of soil organic matter (SOM) (Luo et al. 2015; Pereira et al. 2019; Santos et al. 2018; Yao et al. 2014).
There was a higher total phenolic acid content in the soil in June and September than in March and December in both the pure and mixed plantation stands (Figure 3). This may be caused by higher temperature and more rainfall leading to increased soil microbial activity and in turn accelerated decomposition of litter, which may lead to formation of phenolic acids (Aerts 2006; Dorrepaal et al. 2005; Kirschbaum 1995; Makkonen et al. 2012; Song et al. 2014; Zhang et al. 2008).
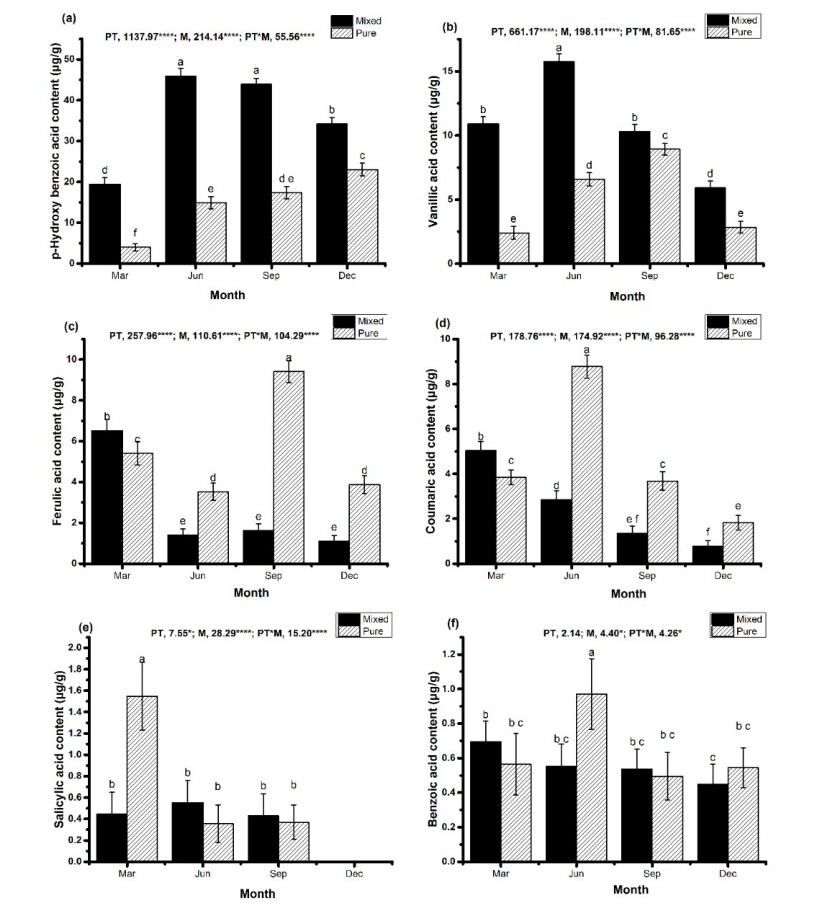
Figure 4: The contents of individual phenolic acids in the soil of the mixed (Eucalyptus grandis × E. urophylla and Erythrophleum fordii Oliv.) and pure (E. grandis × E. urophylla) plantations in different seasons (March, June, September, and December). Data are presented as mean ± SEM. Bars with different lowercase letters indicate significant difference at p < 0.05. F values from the two-way ANOVA analysis are given above each chart. PT, plantation type; M, month; * p < 0.05, *** p < 0.001. / Abbildung 4: Der Gehalt an einzelnen Phenolsäuren in den gemischten (Eucalyptus grandis × E. urophylla und Erythrophleum fordii Oliv.) beziehungsweise reinen (E. grandis × E. urophylla) Bepflanzungen in den verschiedenen Jahreszeiten (März, Juni, September, und Dezember). Daten sind dargestellt als Mittelwert ± SEM. Mit unterschiedlichen Kleinbuchstaben bezeichnete Säulen zeigen signifikante Unterschiede bei p < 0.05. Oberhalb eines jeden Diagrammes sind die F-Werte der zweifachen ANOVA angegeben. PT: Bepflanzungstyp; M: Monat: * p < 0.05, ***: p < 0.001.
3.2. The effect of plantation type on the seasonal dynamics of six phenolic acids in the soil
The p-hydroxybenzoic acid content was the highest in the soil of the mixed plantation. The p-hydroxybenzoic acid content in mixed plantation soils increased significantly (p < 0.05) during June and September compared to March and December, whereas in pure plantation soil, there was a gradual increase in the p-hydroxybenzoic acid content from March through December (Figure 4a). Similarly, the content of vanillic acid was also higher in the mixed plantation than in the pure plantation (Figure 4b). Phenolic acids are the most widely studied and active allelochemicals. Studies have shown that phenolic acids can have an effect (positive or negative) on soil-borne pathogens as well as play an important role in signalling plant–microbe symbiosis (Mandal et al. 2010). Further, phenolic acids can have a hormetic effect on the seed germination or growth of plants (Li et al. 2009; Song et al. 2006). Although the content of p-hydroxybenzoic acid in the soil of the pure plantation and the mixed plantation was high, further experimental investigation is required to reveal whether it is actually the most important allelochemical substance in the soil. In general, the content of ferulic acid, benzoic acid, salicylic acid, and coumaric acid was higher in the pure plantation than in the mixed plantation in most of the studied months (Figure 4c–f). Interestingly, except in the case of benzoic acid, a significant effect (p < 0.001) of plantation type (mixed vs. pure), season, and interaction of season and plantation type was found on the content of each type of phenolic acid in the soil (Figure 4).
The results of the present experiment indicate that the total content of the six phenolic acids in the soil of the mixed plantation was significantly higher than that in the pure plantation, suggesting that mixed planting of E. grandis × E. urophylla and Erythrophleum fordii may increase the content of phenolic acids in the soil. It was observed that while the content of p-hydroxybenzoic acid and vanillic acid was both higher in the mixed plantation soil, the content of the four other phenolic acids was higher in the pure plantation soil. This indicates that the total content of phenolic acids does not necessarily change in the same manner as the content of each single phenolic acid, and the content of one phenolic acid does not necessarily change in the same manner as that of another phenolic acid. The allelopathy of phenolic acids is not only related to the total content, but also to the content of individual phenolic acids (Li et al. 2013; Yang et al. 2017). Therefore, the result suggests that while the effect of total content of phenolic acids is an important consideration in the study of the allelopathic effect of phenolic acids in the soil, the effect of the content of individual phenolic acids should also be considered. The allelopathy of phenolic acids is complex. The allelopathy of some phenolic acids may be stronger at high concentration, while the allelopathy of other phenolic acids may be strong at low concentrations. The allelopathy of some phenolic acids may even fluctuate with increasing concentration (Chen et al. 2016; Gu et al. 2013; Kuiters 1989; Reigosa and Pazos-Malvido 2007; Yang et al. 2010; Yuan et al. 2010). Some phenolic acids may have an effect on one type of plant, but not on another (Reigosa et al. 1999). Some phenolic acids may have no effect or a weak effect when they act alone, while they may have a stronger effect when there is more than one phenolic acid present (Li et al. 2013; Reigosa et al. 1999; Yang et al. 2017). Therefore, further research is needed to determine whether the mixture of Eucalyptus and Erythrophleum fordii can reduce the potential allelopathy of phenolic acids.
3.3. The effect of stand density and understory on phenolic acids
The change in the stand density of a forest may affect the availabilities of light and water by changing light interception, rainfall interception, and transpiration in the forest (Bucci et al. 2008; Chen et al. 2017). Then, the changes in light and water availabilities may affect understory vegetation, soil nutrients, SOM, and soil microorganisms (Bastianelli et al. 2017; Noh et al. 2010; Razafindrabe et al. 2010; Xiang et al. 2014; Zhang et al. 2018; Zhou et al. 2017). The main sources of phenolic acids in soil are the secretion from plant roots (Huang et al. 2016; Jana et al. 2007; Yu and Matsui 1994), the leaching from plant aboveground parts (Hyder et al. 2002), the decomposition of plant litter and SOM (Lü et al. 2012; Zhen et al. 2004). Therefore, the changes in understory vegetation, soil nutrients, SOM, and soil microorganisms may affect phenolic acids in the soil of the forest. In this study, Erythrophleum fordii was introduced into a Eucalyptus plantation as the understory for converting the pure plantation to a multiple-layer forest structure. Consequently, the stand density increased and likely caused changes in the amount of transpiration, available light, and rainfall interception, which might affect the turnover of soil phenolic acids. The total density of the mixed plantation was 1.5 times that of the pure plantation, but the total phenolic acid content of the soil of the mixed plantation in March, June, September, and December was 2.42, 1.91, 1.44, and 1.32 times that of the pure plantation, which is not proportional to the difference in total density. Similar phenomena occurred for the content of individual phenolic acids. The above results show that the effect of the mixed plantation on phenolic acid content in forest soil is not a simple density effect; they indicate the effect of other factors. Thus, the differences in phenolic acid content between the mixed plantation and the pure plantation might primarily be attributed to the introduction of Erythrophleum fordii and the increased total stand density. The interspecific relationship of mixed species and the impact of a multi-layer forest with multiple species on soil processes should be further studied. Future studies on the combined allelopathic effect of phenolic acids on soil properties would be helpful to evaluate the effectiveness of mixed forests.
From the results of the sample plot survey (Table 1), there were no significant differences (p < 0.05) in the DBH or height of Eucalyptus between the pure plantation and the mixed plantation, which may be because the canopy of Eucalyptus in the mixed forest is in the upper layer and the density of Eucalyptus in mixed forest is the same as that in the pure forest. The mortality rates of Eucalyptus in the pure plantation, Eucalyptus in the mixed plantation, and Erythrophleum fordii in the mixed plantation were similar, which may be because the canopy of Eucalyptus in the mixed forest is in the upper layer and the seedlings of Erythrophleum fordii are relatively shade tolerant in young forests (Yi et al. 2018). The interaction between species in a mixed forest is complex (Bouillet et al. 2008; Chauvat et al. 2011; Forrester and Albrecht 2014; Forrester et al. 2010). We cannot yet determine, whether there is allelopathy between Eucalyptus and Erythrophleum fordii in the mixed forest based on our current results. A more in-depth study is needed.
4. Conclusions
Compared with a pure Eucalyptus plantation, the content of each of the six phenolic acids in the soil of the mixed Eucalyptus-Erythrophleum fordii plantation was substantially different over four seasons. In terms of the total content of phenolic acid, mixed planting may result in a greater allelopathic effect of phenolic acids in the soil. For each of the six phenolic acids, mixed planting resulted in increased content of p-hydroxybenzoic acid and vanillic acid in the soil and reduced the content of ferulic acid, coumaric acid, benzoic acid, and salicylic acid in the soil. These findings indicate the need to incorporate both total phenolic acid content and the content of each individual phenolic acid in studies related to the impact of mixed plantations on soil phenolic acid content.
Acknowledgements
This paper was supported by the National Natural Science Foundation of China (No. 31260176), and its financial support is greatly appreciated.
Conflict of interest
The authors declare that they have no conflict of interest.
References
Aerts, R. 2006. The freezer defrosting: global warming and litter decomposition rates in cold biomes. Journal of Ecology 94 (4): 713–724.
Bastianelli, C., Ali, A.A., Beguin, J., Bergeron, Y., Paré, D., Grondin, P., Hély, C. 2017. Boreal coniferous forest density leads to significant variations in soil physical and geochemical properties. Biogeosciences 14 (14): 3445-3459.
Behera, N., Sahani, U. 2003. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. Forest Ecology and Management 174 (1–3): 1–11.
Blum, U. 1996. Allelopathic interactions involving phenolic acids. Journal of Nematology 28 (3): 259–267.
Blum, U., Dalton, B.R. 1985. Effects of ferulic acid, an allelopathic compound, on leaf expansion of cucumber seedlings grown in nutrient culture. Journal of Chemical Ecology 11 (3): 279–301.
Blum, U., Gerig, T.M. 2005. Relationships between phenolic acid concentrations, transpiration, water utilization, leaf area expansion, and uptake of phenolic acids: nutrient culture studies. Journal of Chemical Ecology 31 (8): 1907–1932.
Bolte, M.L., Bowers, J., Crow, W.D., Paton, D.M., Sakurai, A., Takahashi, N., Ujiie, M., Yoshida, S. 1984. Germination inhibitor from Eucalyptus pulverulenta. Agricultural and Biological Chemistry 48 (2): 373–376.
Bouillet, J.P., Laclau, J.P., Gonçalves, J.L.M., Moreira, M.Z., Trivelin, P.C.O., Jourdan, C., Silva, E.V., Piccolo, M.C., Tsai, S.M., Galiana, A. 2008. Mixed-species plantations of Acacia mangium and Eucalyptus grandis in Brazil: 2: Nitrogen accumulation in the stands and biological N2 fixation. Forest Ecology and Management 255(12): 3918-3930.
Bouillet, J.P., Laclau, J.P., Voigtlaender, M., Gava, J.L., Leite, F.P., Hakamada, R., Mareschal, L., Mabiala, A., Tardy, F. 2013. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. Forest Ecology and Management 301(4): 89-101.
Bravo-Oviedo, A., Pretzsch, H., Ammer, C., Andenmatten, E., Barbati, A., Barreiro, S., Brang, P., Bravo, F., Coll, L., Corona, P. 2014. European mixed forests: definition and research perspectives. Forest Systems 23(3): 518-533.
Bucci, S.J., et al. 2008. Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agricultural and Forest Meteorology 148 (6): 839-849.
Carnol, M., Baeten, L., Branquart, E., Grégoire, J.C., Heughebaert, A., Muys, B., Ponette, Q., Verheyen, K. 2014. Ecosystem services of mixed species forest stands and monocultures: comparing practitioners' and scientists' perceptions with formal scientific knowledge. Forestry 87(5): 639-653.
Chauvat, M., Titsch, D., Zaytsev, A.S., Wolters, V. 2011. Changes in soil faunal assemblages during conversion from pure to mixed forest stands. Forest Ecology and Management 262 (3): 317-324.
Chen, E., Zhang, D.H., Wang, D.D., Jin, H., Li, X.Z., He, X.F., Yan, Z.Q., Shen, H.M. 2016. Allelopathic effect and mechanism of action of three phenolic acids on lettuce seedlings. Chinese Journal of Pesticide Science 18(3): 317-322.
Chen, J., Zhao, C.Z., Wang, J.W., Zhao, L.C. 2017. Canopy structure and radiation interception of Salix matsudana: stand density dependent relationships. Chinese Journal of Plant Ecology 41 (6): 661-669.
Chen, Q.B., Peng, L.X., He, L.M. 2002. Allelopathic substances in root and rhizosphere soil of Eucalyptus 12ABL. Chinese Journal of Tropical Crops 22 (4): 28–34.
Dalton, B.R., Blum, U., Weed, S.B. 1989. Differential sorption of exogenously applied ferulic, p-coumaric, p-hydroxybenzoic, and vanillic acids in soil. Soil Science Society of America Journal 53 (3): 757–762.
Dorrepaal, E., Cornelissen, J.H.C., Aerts, R., WallÉN, B.O., van Logtestijn, R.S.P. 2005. Are growth forms consistent predictors of leaf litter quality and decomposability across peatlands along a latitudinal gradient? Journal of Ecology 93 (4): 817–828.
Fang, X., Zhao, L., Zhou, G.Y., Huang, W.J., Liu, J.X. 2015. Increased litter input increases litter decomposition and soil respiration but has minor effects on soil organic carbon in subtropical forests. Plant and Soil 39(1-2): 139-153.
Forrester, D.I., Albrecht, A.T. 2014. Light absorption and light-use efficiency in mixtures of Abies alba and Picea abies along a productivity gradient. Forest Ecology and Management 328: 94-102.
Forrester, D.I., Bauhus, J., Cowie, A.L. 2005. Nutrient cycling in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. Canadian Journal of Forest Research 35(12): 2942-2950+2959.
Forrester, D.I., Theiveyanathan, S., Collopy, J.J., Marcar, N.E. 2010. Enhanced water use efficiency in a mixed Eucalyptus globulus and Acacia mearnsii plantation. Forest Ecology and Management 259(9): 1761-1770.
Gu, Y., Chang, Z.Z., Yu, J.G., Zong, L.G. 2013. Allelopathic effects of exogenous phenolic acids composted by wheat straw on seed germination and seedling growth of rice. Jiangsu Journal of Agricultural Sciences 29(2): 240-246.
Huang, H.J., Liu, C., Wei, S.H., Wang, J.X., Zhang, C.X. 2016. Dynamic root exudation of phenolic allelochemicals from Johnson grass (Sorghum halepense). Weed Biology and Management 15 (4): 133-137.
Hyder, P.W., Fredrickson, E.L., Estell, R.E., Lucero, M.E. 2002. Transport of phenolic compounds from leaf surface of creosotebush and tarbush to soil surface by precipitation. Journal of Chemical Ecology 28 (12): 2475-2482.
Iannucci, A., Fragasso, M., Platani, C., Papa, R. 2013. Plant growth and phenolic compounds in the rhizosphere soil of wild oat (Avena fatua L.). Frontiers in Plant Science 4: 509.
Inderjit. 1996. Plant phenolics in allelopathy. Botanical Review 62 (2): 186–202.
Jagger, P., Pender, J. The role of trees for sustainable management of less favored lands: the case of Eucalyptus in Ethiopia. IFPRI, Washington, USA, EPTD discussion papers 65, 2000.
Jagger, P., Pender, J. 2003. The role of trees for sustainable management of less-favored lands: the case of Eucalyptus in Ethiopia. Forest Policy and Economics 5 (1): 83–95.
Jana, K., Nadezda, V., Jan, T. 2007. Exudation of allelopathic substances in buckwheat (Fagopyrum esculentum Moench). Journal of Agricultural and Food Chemistry 55 (16): 6453-6459.
Kirschbaum, M.U.F. 1995. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage. Soil Biology and Biochemistry 27 (6): 753-760.
Knoke, T., Ammer, C., Stimm, B., Mosandl, R. 2008. Admixing broadleaved to coniferous tree species: a review on yield, ecological stability and economics. European Journal of Forest Research 127(2): 89-101.
Kuiters, A.T., 1989. Effects of phenolic acids on germination and early growth of herbaceous woodland plants. Journal of Chemical Ecology 15(2): 467-479.
Kuiters, A.T. 1990. Role of phenolic substances from decomposing forest litter in plant-soil interactions. Acta Botanica Neerlandica 39 (4): 329–348.
Kuiters, A.T., Sarink, H.M. 1986. Leaching of phenolic compounds from leaf and needle litter of several deciduous and coniferous trees. Soil Biology and Biochemistry 18 (5): 475–480.
Laclau, J.P., Ranger, J., de Moraes Gonçalves, J.L., Maquère, V., Krusche, A.V., M’Bou, A.T., Nouvellon, Y., et al. 2010. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations. Forest Ecology and Management 259 (9): 1771–1785.
Li, X., Zhou, B.L., Chen, S.L., Lin, S.S. 2009. Effects of the autotoxic substance in eggplants on pepper seed germination and Fusarium oxysporum f. sp. Acta Ecologica Sinica 29 (2): 960–965.
Li, Z.L., Hui, Z.L., Zhang, J.L., Shen, B.Y., Li, C.Z. 2013. Study on the effects and mechanism of exogenous phenolic acids on potato's growth and development. Acta Agriculturae Boreali-Sinica 28(6): 147-152.
Liu, Y., Zhang, J., Feng, M.S. 2006. Dynamics of litter production, nutrient return and decomposition of four Eucalyptus grandis plantation. Scientia Silvae Sinicae 42(7): 1-10.
Luo, D., Shi, Z.M., Wang, W.X., Liu, S.R., Lu, L.H., Ming, A.G., Yu, H.L. 2015. Carbon and nitrogen storage in monoculture and mixed young plantation stands of Erythrophleum fordii and Pinus massoniana in subtropical China. Acta Ecologica Sinica 35 (18): 6051-6059.
Lü, W.G., Yang, G.C., Liu, L., Zhang, C.L., Shen, Q.R., Li, S.X., Zheng, X.Q. 2012. Phenolic acids in decomposing residuces of watermelon. Acta Agriculturae Boreali-Sinica 27 (S1): 154-157.
Makkonen, M., Berg, M.P., Handa, I.T., Hättenschwiler, S., Ruijven, Jv., Bodegom, P.Mv., Aerts, R. 2012. Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecology Letters 15 (9): 1033–1041.
Mandal, S.M., Chakraborty, D., Dey, S. 2010. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signaling and Behavior 5 (4): 359–368.
Martens, D.A. 2002. Relationship between plant phenolic acids released during soil mineralization and aggregate stabilization. Soil Science Society of America Journal 66 (6): 1857–1867.
Mason, W.L., Connolly, T. 2014. Mixtures with spruce species can be more productive than monocultures: evidence from the Gisburn experiment in Britain. Forestry 87(2): 209-217.
Meng, M., Li, J., Li, L.F., Liu, Y.G., Chen, H.W., Feng, X. 2011. Litter amounts and decomposition dynamics of Pinus kesiya var.langbianensis plantation. Journal of West China Forestry Science 40(2): 56-63.
Noh, N.J., et al. 2010. Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. Journal of Plant Research 123 (4): 411-419.
Patrick, Z.A. 1971. Phytotoxic substances associated with the decomposition in soil of plant residues. Soil Science 111 (1): 13–18.
Patterson, D.T. 1981. Effects of allelopathic chemicals on growth and physiological responses of soybean (Glycine max). Weed Science 29 (1): 53–59.
Pereira, A.P.A., Durrer, A., Gumiere, T., Gonçalves, J.L.M., Robin, A., Bouillet, J.-P., Wang, J.T., Verma, J.P., Singh, B.K., Cardoso, E.J.B.N. 2019. Mixed Eucalyptus plantations induce changes in microbial communities and increase biological functions in the soil and litter layers. Forest Ecology and Management 433: 332-342.
Razafindrabe, B.H.N., He, B., Inoue, S., Ezaki, T., Shaw, R. 2010. The role of forest stand density in controlling soil erosion: implications to sediment-related disasters in Japan. Environmental Monitoring and Assessment 160 (1-4): 337.
Reigosa, M.J., Pazos-Malvido, E. 2007. Phytotoxic effects of 21 plant secondary metabolites on Arabidopsis thaliana germination and root growth. Journal of Chemical Ecology 33(7): 1456-1466.
Reigosa, M.J., Souto, X.C., Gonz´Lez, L. 1999. Effect of phenolic compounds on the germination of six weeds species. Plant Growth Regulation 28(2): 83-88.
Santos, F.M., Balieiro, F.d.C., Fontes, M.A., Chaer, G.M. 2018. Understanding the enhanced litter decomposition of mixed-species plantations of Eucalyptus and Acacia mangium. Plant and Soil 423(1-2): 141-155.
Song, L., Pan, K.W., Wang, J.C., Ma, Y.H. 2006. Effects of phenolic acids on seed germination and seedling antioxidant enzyme activity of alfalfa. Acta Ecologica Sinica 26 (10): 3393–3403.
Song, Y., Gu, X.R., Yan, H.Y., Mao, W.T., Wu, X.L., Wan, Y.X. 2014. Dynamics of microbes and enzyme activities during litter decomposition of Pinus massoniana forest in mid-subtropical area. Environmental Science 35 (3): 1151–1158.
Tan, X.M., Wang, H.T., Kong, L.G., Wang, Y.P. 2008. Accumulation of phenolic acids in soil of a continuous cropping Poplar plantation and their effects on soil microbes. Journal of Shandong University 43(1): 14-19.
The State Forestry Administration of the People’s Republic of China. 2014. China Forestry Resource Report. China Forestry Publishing House, Beijing, p 75.
Wang, H., Liu, S.R., Wang, J.X., Shi, Z.M., Lu, L.H., Zeng, J., Ming, A.G., Tang, J.X., Yu, H.L. 2013. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. Forest Ecology and Management 300: 4–13.
Wang, H.G., Zhang, J., Yang, W.S., Huang, Q.M., Zou, P. 2006. A research on the allelopathic substances in root system and root system soil of Eucalyptus grandis. Journal of Sichuan Normal University (Natural Science) 29 (3): 368.
Wang, J.G., Zhang, J., Li, X.W. 2007. The study on spatial distribution of allelochemicals in the soil of Eucalyptus grandis plantation. Journal of Sichuan Agricultural University 25 (2): 121–126.
Wang, Q.K., Wang, S.L. 2008. Soil microbial properties and nutrients in pure and mixed Chinese fir plantations. Journal of Forest Research 19 (2): 131–135.
Wang, W.X., Shi, Z.M., Luo, D., Liu, S.R. 2016. Decomposition characteristics of leaf litter and fine roots of Erythrophleum fordii and Castanopsis hystrix in subtropical China. Acta Ecologica Sinica 36 (12): 3479-3487.
Wang, Y.P. 2010. Degradation mechanism of continuous cropping poplar plantations: accumulation and allelopathy of phenolic acids. Shandong Agricultural Univerisity, Shandong.
Wen, Y.G. 2005. The ecological problems and sustainable management of industrial Eucalyptus plantation. Journal of Guangxi Academy of Sciences 21 (1): 13–19.
Wolf, D.C., Dao, T.H., Scott, H.D., Lavy, T.L. 1989. Influence of sterilization methods on selected soil microbiological, physical, and chemical properties. Journal of Environmental Quality 18(1): 39-44.
Xavier, M., Lorenz, F., Michael, S.L., Harald, B. 2011. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecology Letters 14(12): 1211-1219.
Xiang, Y.B., Huang, C.D., Hu, T.X., Tu, L.H., Li, R.H., Yan, Z., Pu, M. 2014. Characteristics of soil organic carbon and microbial biomass carbon and nitrogen in Eucalyptus grandis plantations with different densities. Acta Botanica Boreali-Occidentalia Sinica 34 (7):1476-1481.
Xing, W., Lu, X.M., Xu, F.W., Ying, J.Y., Chen, D.M., Bai, Y.F. 2019. Linking microbial community structure to carbon substrate chemistry in soils following aboveground and belowground litter additions. Applied Soil Ecology 141: 18-25.
Yang, X.Y., Li, L., Zhang, Q., Xie, K., Li, J.Y., He, H.B. 2017. Allelopathy of phenolic acids with different types on the growth of lettuce (Lactuca sativa L.). Journal of Fujian Agriculture and Forestry University 46(1): 21-26.
Yang, Y., Wang, H.T., Wang, Y.P., Jiang, Y.Z., Wang, Z.Q. 2010. Effects of exogenous phenolic acids on root physiologic characteristics and morphologic development of poplar hydroponic cuttings. Scientia Silvae Sinicae 46(11): 73-80.
Yao, Y., Wang, R., Lu, J.K., Sui, X.H., Wang, E.T., Chen, W.X. 2014. Genetic diversity and evolution of Bradyrhizobium populations nodulating Erythrophleum fordii, an evergreen tree indigenous to the southern subtropical region of China. Applied and Environmental Microbiology 80 (19): 6184-6194.
Ye, S.F., Zhou, Y.H., Sun, Y., Zou, L.Y., Yu, J.Q., 2006. Cinnamic acid causes oxidative stress in cucumber roots, and promotes incidence of Fusarium wilt. Environmental and Experimental Botany 56(3): 255-262.
Yi, W.J., Zhang, H.D., Ye, S.M., Yang, M. 2018. The effects of light intensity on seedling growth and photosynthetic characteristics of Erythrophleum fordii. South China Forestry Science 46(1): 29-32+37.
Yu, J.Q., Matsui, Y. 1994. Phytotoxic substances in root exudates of cucumber (Cucumis sativus L.). Journal of Chemical Ecology 20 (1): 21-31.
Yuan, N., Zheng, R.H., Gu, X.P., Wu, X.L., Yue, J.J. 2010. Allelopathic effect of phenolic acids on Phyllostachys edulis seedlings. Journal o f Nanjing Forestry University 34(1): 29-33.
Zeng, R.S., Li, P.W. 1997. Allelopathic effects of Eucalyptus exserta and E. urophylla. Journal of South China Agricultural University 18 (1): 6–10.
Zhai, M.P., Jia, L.M. 1993. Allelopathy of forest plants. Journal of Beijing Forestry University 15 (3): 138–147.
Zhang, D.Q., Hui, D.F., Luo, Y.Q., Zhou, G.Y. 2008. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. Journal of Plant Ecology 1 (2): 85-93.
Zhang, L.H., Qi, J.Q., Liu, P.Y., Zhou, K.L., Li, X., Li, Q., Hao, J.F. 2018. Effects of stand density on community structure and species diversity of Eucalytus robusta plantation. Acta Botanica Boreali-Occidentalia Sinica 38 (1): 166-175.
Zhen, W.C., Wang, X.Y., Kong, J.Y., Cao, K.Q. 2004. Determination of phenolic acids in root exudates and decomposing products of strawberry and their allelopathy. Journal of Agricultural University of Hebei 27 (4): 74-78.
Zhou, S.P., Liang, K.N., Du, J., Li, B.J., Zhou, Z.Z., Huag, G.H. 2017. Research on understory vegetation and soil physical-chemical properties of teak plantation with difference stand densities. Bulletin of Botanical Research 37 (2): 200-210.
Zhou, X., Yu, G., Wu, F. 2012. Soil phenolics in a continuously mono-cropped cucumber (Cucumis sativus L.) system and their effects on cucumber seedling growth and soil microbial communities. European Journal of Soil Science 63(3): 332-340.
Zhu, L.X., Zhang, J.E., Liu, W.G. 2003. Review of studies on interactions between root exudates and rhizopheric microorganisms. Frontiers in Ecology and the Environment 12 (1): 102–105.



