Schlüsselbegriffe: Nachbaranalyse, räumliche Verteilung, Konkurrenz, generalisierte lineare Modelle, Picea abies, Fagus sylvatica, Pinus brutia
Abstract
The competition of individual trees for natural resources results in growth of their height as well as their basal area. While height growth is primarily driven by environmental conditions, growth of basal area appears limited by growing space. We adopted the spatial neighborhood technique in this study to determine the effect that competition has on tree basal area in three forests in Greece: a) mixed Norway spruce (Picea abies) forest, b) pure European beech (Fagus sylvatica) forest, and c) a mixed Turkish pine (Pinus brutia) forest. We randomly selected in total 337 sample (target) trees. Using the Generalized Linear Modeling (GLM) method, basal area at breast height of target trees were modelled based on stump height (20 cm above the ground) diameter and distance to neighboring trees and the species of the neighbors. The distance between neighboring and target trees and species of the neighbors substantially improved the model performance for the mixed pine and spruce forests, while considering neighbors led to less precise predictions for the pure beech forest. Our study suggests, that the selected pure and mixed forests have a different response to competition. Apparently, the species of the competing trees and the combination of distance and species of neighbors have a more complicated effect on the competition on trees than previously expected.
Zusammenfassung
Die Konkurrenz der einzelnen Bäume um natürliche Ressourcen führt zum Wachstum ihrer Höhe und ihrer Grundfläche. Während das Höhenwachstum in erster Linie von den Umweltbedingungen bestimmt wird, scheint das Wachstum der Grundfläche durch den verfügbaren Platz begrenzt zu sein. Wir haben die räumliche Nachbaranalyse in dieser Studie verwendet, um zu untersuchen, welchen Effekt Konkurrenz auf die Baumgrundfläche in drei Wäldern in Griechenland hat: a) gemischter Fichtenwald (Picea abies), b) reiner Rotbuchenwald (Fagus sylvatica) und c) gemischter Kiefernwald (Pinus brutia). Wir haben insgesamt 337 Probebäume (Zielbäume) zufällig ausgewählt. Mit der Generalized Linear Modeling (GLM)-Methode wurde die Grundfläche in Brusthöhe von Zielbäumen mittels Stumpfhöhe (20 cm über dem Boden) sowie dem Durchmesser und Abstand benachbarter Bäume und deren Baumart modelliert. Der Abstand zwischen Nachbar- und Zielbäumen und den Arten der Nachbarn verbesserte die Modellleistung für die Kiefern- und Fichtenmischwälder erheblich, während die Berücksichtigung der Nachbarn zu weniger genauen Vorhersagen für den reinen Buchenwald führte. Unsere Studie zeigt, dass die untersuchten Rein- und Mischwälder eine unterschiedliche Reaktion auf Konkurrenz aufweisen. Scheinbar haben die Arten der konkurrierenden Bäume und die Kombination von Entfernung und Arten der Nachbarn einen komplexeren Einfluss auf die Konkurrenz an Bäumen als bisher erwartet.
1. Introduction
Plants are well known for the plasticity of their growth in response to competition (Jack and Long 1991, Weiner and Thomas 1992, Takahashi 1996, Muth and Bazzaz 2003, Vincent and Harja 2007, Thorpe et al. 2010,). The structure of a plant community is determined by competitive interactions among members (Barnes and Spurr 1998, Harper 2010). It is very important for ecosystem management and forest ecology to understand, how trees interact. Latest changes in forest management goals have resulted into an increased interest in maintaining and/or increasing compositional and structural complexity in managed forests (Burton 2003, Kohm et al. 1997, Puettmann et al. 2014). While traditionally specific and even-aged forestry practices were more common, there is now more emphasis on mixed species management using different spatial indices considering variation in tree dimension and their spatial distribution (Bauhus et al. 2009, Harvey et al. 2002, Kuuluvainen 2009, Canham and Uriarte 2006). Thus, the prediction of stand development in complex forests require a better understanding of tree size, species, and neighborhood competition.
This study used estimates of basal area at breast height, considering the possible spatial interaction of individual trees using the distances between them (Canham and Uriarte 2006, Gratzer et al. 2004, Stoll et al. 1994). Previous studies using neighborhood analysis analysed annual plant growth (Watkinson et al. 1983, Weiner and Thomas 1992), adult tree growth (Coates et al. 2009, Papaik and Canham 2006, Uriarte et al. 2004a, Canham et al. 2004), sapling and seedling demography (Uriarte et al. 2004b), tree mortality (Thorpe et al. 2008, Uriarte et al. 2004a), and tree crown size (Thorpe et al. 2010) as elements of immediate “neighborhood” conditions, which include the size, species, distance, and density of neighbors. Stand density measures such as basal area per hectare or “Area Potentially Available” (APA) (Daniels et al. 1986) are distance-dependent metrics of competition for tree growth. However, stand density does not reflect the arrangement and clustering of individual trees, since the distance of neighbor trees influences assymetrically their growth potential (Tomé and Burkhart 1989). Mathematical formulas were derived to express the effect of neighbors on individual tree growth (Burkhart and Tomé 2012). Other models consider more complex competition indices taking into account the size, distance and number of neighbors (Weiskittel et al. 2011). Nevertheless, the effect of neighbor characteristics, such as species and their distance, has not been studied in the context of individual tree growth modeling, except as in the context of stand density.
The aim of this study is to explore the influence of clustering on basal area using distances and species neighboring trees for three species, Norway spruce (Picea abies), European beech (Fagus sylvatica), and Turkish pine (Pinus brutia) in Greece.
2. Materials and methods
2.1 Study site and field sampling
Three study sites in Northern Greece were selected; Elatia (Karantere) (Picea abies was the studied species), Kastaneri - Livadia (Fagus sylvatica was the studied species), and Kedrinos Lofos (Seich Sou) (Pinus brutia was the studied species). Random point sampling of dominant trees was used (Diggle et al. 1976). Dominant trees are defined as the trees having crowns extending above the general level of the main canopy and receiving full light from above and partly from the sides (Oliver and Larson 1996).
Ninety-six (96) dominant Norway spruce trees were selected from the mixed forest of Elatia (Karantere) that extends along the Greek – Βulgarian border. Altitude ranges from 1480 to 1610 m. The soil is sandy – clay and the parent rock is granite – gneiss (Lepidi 2010). Mean annual rainfall is 694 mm and mean average temperature is 11.4 °C at the nearest weather station at 600 m altitude (Grigoriadis et al. 2007).
One hundred (100) dominant European beech trees were selected from the pure beech forest of Kastaneri – Livadia, on the eastern side of Mount Paiko. Altitude ranges from 1160 m to 1440 m. The soil is sandy – clay and the parent rock is shale and limestone (Vourtsas et al. 2014), while the mean annual rainfall is 628 mm and the mean average temperature is 10.4 °C (Zerva et al. 2008).
One hundred forty-one (141) dominant Turkish pine trees were selected from a mixed part of the peri-urban forest of Kedrinos Lofos (Seich Sou), in the north-northeast of the city of Thessaloniki. The peri-urban forest is a planted forest consisted mainly of Turkish pine trees. Altitude ranges from 170 to 470 m. The soil is sandy – clay and the parent rock is gneiss, calcareous mica schists and quartzite, while the mean annual rainfall is 447.8 mm and the mean average temperature is 15.8 °C (Gatzogiannis et al. 1996; Andreadis 2005).
Distance was measured from the center of the target tree's basal area (the center of the circular basal area at stump height) to the center of its neigbor's basal area (also the center of a circle), in order to weight the effect of the distance between two trees with different basal areas.
2.2 Data analysis
To determine the effect of competition on basal area at breast height, basal area is estimated via stump height diameter (20 cm above the ground), and via neighborhood parameters and stump height diameter. We used a neighborhood technique to analyze the effect of competition on basal area, by summing up the distances from the three nearest trees to each sampled tree, into a Generalized Linear Model (GLM). The number of the nearest trees (three) was chosen arbitrarily.
The simplest of the tested models, the “stump only” model, uses the stump height diameter as the only independent variable for basal area. The “stump only” model suggests that the basal area at breast height of a specified tree, has positive linear dependence with its stump height diameter. The tested model is:

where BÂ is the estimated basal area at breast height (m2), dst is the stump height diameter (20 cm above the ground), and bi are the linear regression coefficients.
The model of “stump and neighbors” makes an addition to the GLM; it inserts the distances from the three nearest trees, as auxiliaries. In the model of “stump and neighbors”, two scientific hypotheses are identified, regarding the way that neighborhood competition affects tree size. The simplest of the tested hypotheses, the “equivalent competitors” model, suggests that the competitive neighboring effects on the target tree do not differ among species. The model tested is:
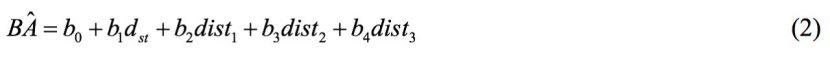
where dist1, dist2, dist3 is the distance of the first closest tree to a target tree, the second closest tree to a target tree, and the third closest tree to a target tree, respectively.
Given the fact that several studies have emphasized with strong evidence of differences that exists in form of species-specific competitive effects on specific tree growth (Coates et al. 2009, Uriarte et al. 2004a, Canham et al. 2004), we examined the effect of neighbor species to the basal area of a target tree, by applying the “full model":
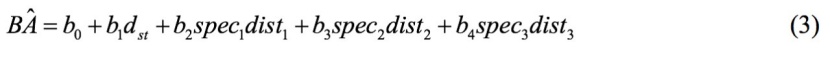
where spec1, spec2, spec3 is the species of the first closest tree to a target tree, the second closest tree to a target tree, and the third closest tree to a target tree, respectively.
In the spruce dominated forest, the target trees were Norway spruce and the neighbors were Scots pine (speci = 1, i = 1,2,3) and Norway spruce (speci = 2, i = 1,2,3); in the beech dominated forest, the target trees were European beech and the neighbors were also beech (pure forest); in the forest dominated by pine, the target trees were Turkish pine and the neighbors were Mediterranean cypress (speci = 1, i = 1,2,3) and Turkish pine (speci = 2, i = 1,2,3). The statistical summary for all measured variables are given in Table 1. The study areas are shown in Figure 1, while photos from the studied forests are shown in Figure 2.
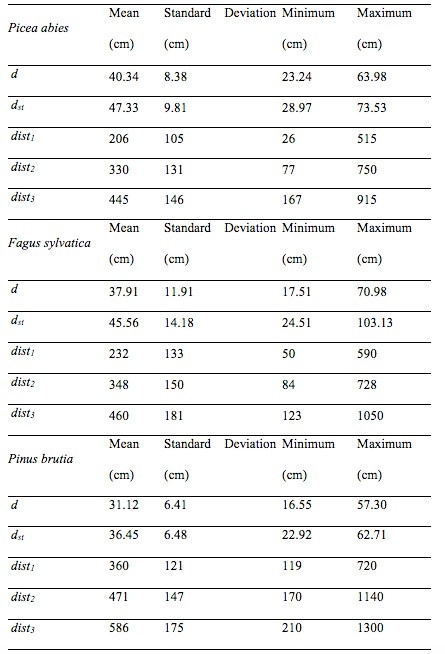
Table 1: Summary statistics of the sampled trees. / Tabelle 1: Zusammenfassende Statistiken zu den untersuchten Bäumen.
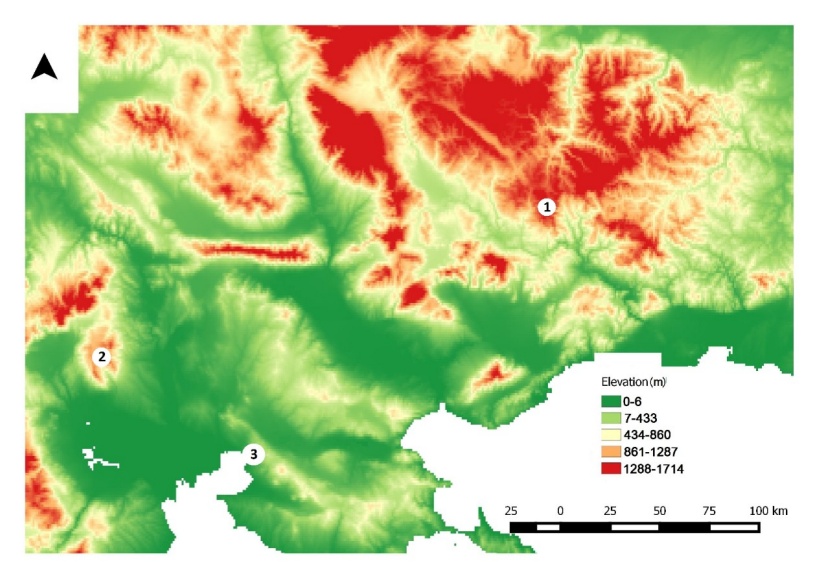
Figure 1: Study areas of the three studied species (1: Elatia (Karantere) for Picea abies; 2: Kastaneri - Livadia for Fagus sylvatica; 3: Kedrinos Lofos (Seich Sou) for Pinus brutia). / Abbildung 1: Untersuchungsgebiete der drei untersuchten Arten (1: Elatia (Karanteren) für Picea abies; 2: Kastaneri - Livadia für Fagus sylvatica; 3: Kedrinos Lofos (Seich Sou) für Pinus brutia).

Figure 2: Photos of the three studied forests (1: Elatia (Karantere) for Picea abies; 2: Kastaneri - Livadia for Fagus sylvatica; 3: Kedrinos Lofos (Seich Sou) for Pinus brutia). / Abbildung 2: Fotos der drei untersuchten Wälder (1: Elatia (Karanteren) für Picea abies; 2: Kastaneri - Livadia für Fagus sylvatica; 3: Kedrinos Lofos (Seich Sou) für Pinus brutia).
2.2.1 Estimation of parameters and selection of model
The Generalized Linear Modeling method was used to fit the candidate models to the data, using the SPSS v.21 software (IBM SPSS Statistics for Windows 2012). The general linear model is expanded by the Generalized Linear Models procedure so that the dependent variable (basal area) is related linearly to the factors (species) and covariates (stump height diameter, distances of neighbor trees) (Özçelík et al. 2010, Thorpe et al. 2010). Also, the dependent variable is allowed to have a distribution that is not normal (in our case, the Kolmogorov-Smirnov test had a p-value of 0.011 < 0.05 for Norway spruce, 0.015 < 0.05 for European beech, and 0.012 < 0.05 for Turkish pine, respectively). The models’ fit was assessed using five statistical comparison criteria (Kitikidou 2005):
1. Absolute Mean Error (AME)
2. Standard Error of the Estimate (SEE)
3. Coefficient of determination R2
4. Root of the Mean Squared Error (RMSE)
5. Sum of Squared Errors (SSE)
We plotted observed basal area values on the Y-axis versus the predicted values on the X-axis to assess model bias (an unbiased model has R2 = 1).
3. Results
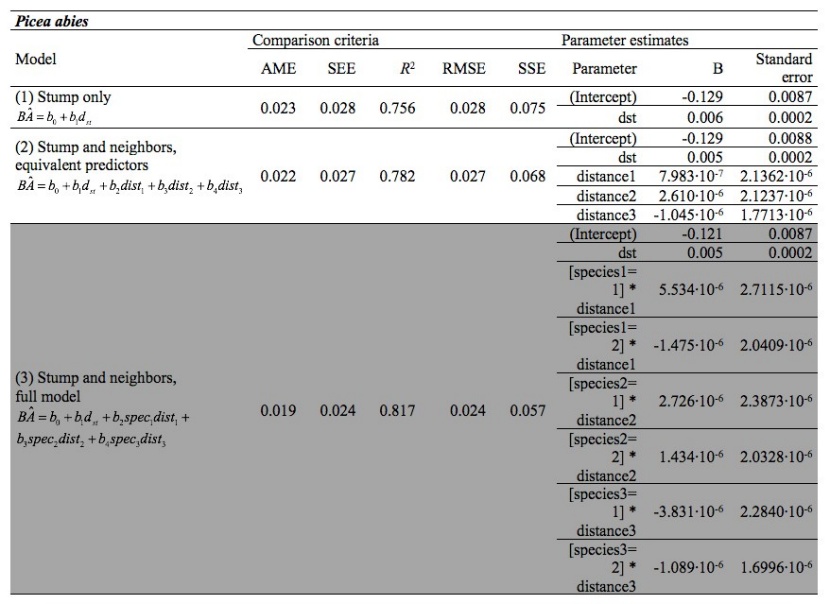
Table 2: Comparison criteria and parameter estimates for Picea abies (selected model is highlighted). / Tabelle 2: Vergleichskriterien und Parameterschätzungen für Picea abies (das ausgewählte Modell ist hervorgehoben).
3.1 Selection of the most well-fitted model
The full model considering stump height diameter, distance and species of neighbors was the most efficient model for estimating basal area for Norway spruce and Turkish pine (Tables 2 and 4). In this model, distance and species of neighbors has explanatory effect on basal area and thus competition. For the pure beech forest, where neighborhood species were not applicable, the distances between the three nearest trees and the target tree did not result in improvement of the basal area estimates (Table 3). Parameter estimates and their standard errors are given in the Tables 2, 3, and 4, for Norway spruce, European beech, and Turkish pine, respectively.
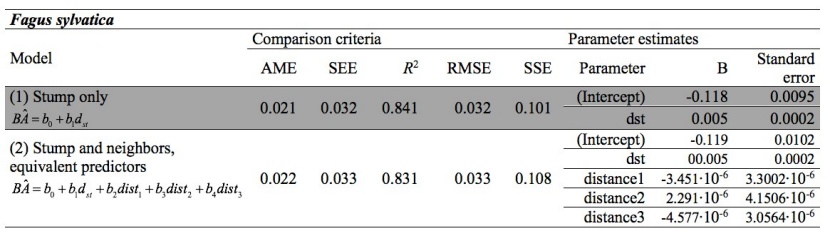
Table 3: Comparison criteria and parameter estimates for Fagus sylvatica (selected model is highlighted). / Tabelle 3: Vergleichskriterien für Fagus sylvatica (das ausgewählte Modell ist hervorgehoben).
4. Discussion
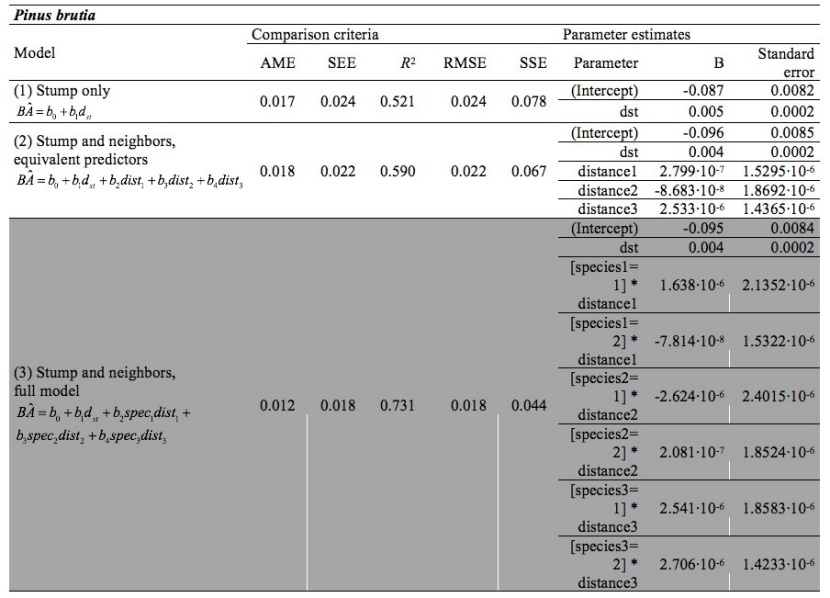
Table 4: Comparison criteria and parameter estimates for Pinus brutia (selected model is highlighted). / Tabelle 4: Vergleichskriterien und Parameterschätzungen für Pinus brutia (das ausgewählte Modell ist hervorgehoben).
Competition for light is an important characteristic of forest ecosystems (Oliver and Larson 1996). This study provides novel insights into how competition affects dominant tree growth for three important and well-known species in Greece. We provide evidence for similarities in species’ response to neighborhood competition. Except for the monospecific beech forest, both identity and distance of neighboring species influenced competitive effects on target trees. Yet, response to crowding from neighbors did not reduce with increasing neighbor tree distance as previously shown (Thorpe et al. 2010). The B values in Tables 3.1 and 3.3 for the full model did not decrease as neighbor distances increase but also, in some cases, the B values were negative. This means that there is no reduction in the effective neighborhood competition when neighbor distance increases. If the effect of the neighbor distance (independent variable) on a target tree decreased with distance, the B value should be larger for the distance of the first neighbor, smaller for the distance of the second neighbor, and smallest for the third (most distant) neighbor. As described in the Introduction, the neighbor effect has been studied in stand models, not individual tree models, so, there are no models in literature to compare with this one; this could be a subject for further research. Possibly the species characteristics of the competing trees, or the combination of distance and species of the competing trees, have a more complicated effect on the competition imposed on the target tree than expected.
The relationship basal area at breast height and stump height diameter is strongly influenced by the taper of the bole, and thus from the competition regime under which a tree grew during its life (Philip 2002, Milios and Akritidou 2002, Kitikidou et al. 2014). Based on the results of the present study, there are evidences that the effect of competition on tree growth is species depended. This means that the effect of competition on the growth of the target tree bole is influenced by growth ecology of the species in which the target tree belongs, and the species of the trees that impose competition on the target tree. In the central Rhodope mountains of northeastern Greece, the growth of dominant beech trees was higher when they competed with Scots pine (Pinus sylvestris) trees, compared with the growth they exhibited when the competitors were beech trees (Milios 2004).
The dominant trees, defined as the trees having crowns extending above the general level of the main canopy, and receiving full light from above and partly from the sides, are the most competitive trees locally in a small group of trees (Oliver and Larson 1996, Smith et al. 1997). Seems that in the case of beech, the dominant trees are, so much taller than the neighbooring trees, that the distance of the closest trees does not influence the competition imposed on them (dominant trees). On the other hand, in spruce and in pine, the information provided by the distances and the species of the closest trees improved the estimation of the basal area, compared to the stump only models (Tables 3.1, 3.3). In the case of pine, the improvement is noticeable. The fact that the B values change as distances increase (Tables 2 and 4) in a very different way between different species, means that different species have different influences upon each other. In an attempt to explain the differences among species, based on their tolerance in shadow, in the pure forest of the shade tolerant species of beech (Korakis 2015), which is characterized by growth plasticity (Milios and Akritidou 2002, Assmann 1970), the form of bole of dominant trees is not affected by the distance of the closest neighboring trees. In the semi-shade tolerant Norway spruce (Korakis 2015), the distance and the species of the closest neighboring trees influence growth of the tree bole. Finally, in the light demanding species of Turkish pine (Korakis 2015), the effect of the distance and species of the neighboring trees on the estimation of the basal area using the stump height diameter is high. Other potential explanations for observed patterns could be the effect of altitude, or the effect of harvesting intensity; these factors could be examined in a follow-up study.
5. Conclusions
In this study, we explored the influence of neighboring trees on basal area at breast height, by recording neighbors' species' and distances. There is evidence that neighborhood competition affect mixed forests, but has little effect in pure forests. More research is needed to specify the influence of distance and species of the competing neighbors on the competition imposed on dominant trees and on trees with other social status as well as other tree species. Also, estimating basal area from stump height diameter following harvesting, taking into account the removed trees in increment calculations, and studying the potential effect of site conditions across different altitudes, would promote research on this topic.
References
Andreadis, K. 2005. Final study of the completion and improvement of the vegetation of the peri-urban forest of Thessaloniki. Reforestation Department of Thessaloniki.
Assmann, E. 1970. The principles of forest yield study. Pergamon Press, Oxford.
Barnes, B., and S. Spurr. 1998. Forest ecology. J. Wiley, New York.
Bauhus, J., K. Puettmann, and C. Messier. 2009. Silviculture for old-growth attributes. Forest Ecology and Management. 258(4):525-537.
Burkhart, H., and M. Tomé, M. 2012. Modeling forest trees and stands. Springer, New York.
Burton, P. 2003. Towards sustainable management of the boreal forest. NRC Research Press, Ottawa.
Canham, C., and M. Uriarte. 2006. Analysis of Neighborhood Dynamics Of Forest Ecosystems Using Likelihood Methods And Modeling. Ecological Applications. 16(1):62-73.
Canham, C., P. LePage, and K. Coates. 2004. A neighborhood analysis of canopy tree competition: effects of shading versus crowding. Canadian Journal of Forest Research. 34(4):778-787.
Coates, K., C. Canham, and P. LePage. 2009. Above- versus below-ground competitive effects and responses of a guild of temperate tree species. Journal of Ecology. 97(1):118-130.
Daniels, R., H. Burkhart, and T. Clason. 1986. A comparison of competition measures for predicting growth of loblolly pine trees. Canadian Journal of Forest Research. 16(6):1230-1237.
DeYoung, J. 2000. Forest Measurements: An Applied Approach. Springer, New York.
Diggle, P., J. Besag, and J. Gleaves, J. 1976. Statistical Analysis of Spatial Point Patterns by Means of Distance Methods. Biometrics. 32(3):659-667.
Gatzogiannis, S., P. Kyriakidis, and C. Gigis. 1996. Management plan of the peri-urban forest of Thessaloniki. Forest Service of Thessaloniki and Institute of Forest Research.
Gratzer, G., C. Canham, U. Dieckmann, A. Fischer, Y. Iwasa, R. Law, and M. Lexer et al. 2004. Spatio-temporal development of forests – current trends in field methods and models. Oikos. 107(1):3-15.
Grigoriadis, N., G. Spiroglou, and J. Petermann. 2007. Chraracteristics of the protected area (GR1140003) of the Elatia Forest at Drama Prefecture region. P. 80–90 in 13th conference of Hellenic forestry society.
Harper, J. 2010. Population biology of plants. Blackburn Press, Caldwell, N.J.
Harvey, B., A. Leduc, S. Gauthier, and Y. Bergeron. 2002. Stand-landscape integration in natural disturbance-based management of the southern boreal forest. Forest Ecology and Management. 155(1-3):369-385.
IBM SPSS Statistics for Windows. 2012. IBM Corp.
Jack, S., and J. Long. 1991. Analysis of stand density effects on canopy structure: a conceptual approach. Trees. 5(1).
Kohm, K., J. Franklin, and J. Thomas. 1997. Creating a forestry for the 21st century. Island Press, Washington D.C.
Korakis, G. 2015. Forest Botany. Hellenic Academic Libraries Link, Athens.
Kuuluvainen, T. 2009. Forest Management and Biodiversity Conservation Based on Natural Ecosystem Dynamics in Northern Europe: The Complexity Challenge. AMBIO: A Journal of the Human Environment. 38(6):309-315.
Kitikidou, K. 2005. Applied statistics with use of the SPSS statistical package. Tziola Publications, Thessaloniki, Greece.
Kitikidou, K., E. Milios, and I. Lipiridis. 2014. Tree volume model estimates and nearest neighbor analysis in the stands of Scots pine (Pinus sylvestris L.) in the central part of Rodope Mountain. Šumarski list. 138:11-12.
Lepidi, A. 2010. Study for the Protection and Management of the Public Forest Complex of the Western Nestos Prefecture of Drama, for the period 2007-2016. Drama Forest Service.
Milios, E. 2004. The influence of stand development process on the height and volume growth of dominant Fagus sylvatica L. s.l. trees in the central Rhodope Mountains of north-eastern Greece. Forestry. 77(1):17-26.
Milios, E., and S. Akritidou. 2002. Growth patterns of beech (Fagus sylvatica s.l.) trees under different growth conditions. P. 676–684 in 10th conference of Hellenic forestry society.
Muth, C., and F. Bazzaz. 2003. Tree canopy displacement and neighborhood interactions. Canadian Journal of Forest Research. 33(7):1323-1330.
Oliver, C., and B. Larson. 1996. Forest stand dynamics. J. Wiley, New York.
Özçelík, R., J. Brooks, M. Diamantopoulou, and H. Wiant, H. (2010). Estimating breast height diameter and volume from stump diameter for three economically important species in Turkey. Scandinavian Journal of Forest Research. 25(1): 32-45.
Papaik, M., and C. Canham. 2006. Multi-model analysis of tree competiton along environmental gradients in southern New England Forests. Ecological Applications. 16(5):1880-1892.
Philip, M. 2002. Measuring trees and forests. CABI, Wallingford [etc.].
Puettmann, K., K. Coates, and C. Messier. 2014. Critique of Silviculture. Island Press, Washington.
Smith, D., B. Larson, M. Kelty, and P. Ashton. 1997. The practice of silviculture. J. Wiley, New York.
Stoll, P., J. Weiner, and B. Schmid. 1994. Growth Variation in a Naturally Established Population of Pinus Sylvestris. Ecology. 75(3):660-670.
Takahashi, K. 1996. Plastic Response of Crown Architecture to Crowding in Understorey Trees of Two Co-dominating Conifers. Annals of Botany. 77(2):159-164.
Thorpe, H., S. Thomas, and J. Caspersen. 2008. Tree mortality following partial harvests is determined by skidding proximity. Ecological Applications. 18(7):1652-1663.
Thorpe, H., R. Astrup, A. Trowbridge, and K. Coates. 2010. Competition and tree crowns: A neighborhood analysis of three boreal tree species. Forest Ecology and Management. 259(8):1586-1596.
Tomé, M., and H. Burkhart. 1989. Distance-dependent competition measures for predicting growth of individual trees. Forest Science. 35(3):816–831.
Uriarte, M., C. Canham, J. Thompson, and J. Zimmerman. 2004a. A neighborhood analysis of tree growth and survival in a hurricane-driven tropical forest. Ecological Monographs. 74(4):591-614.
Uriarte, M., R. Condit, C. Canham, and S. Hubbell. 2004b. A spatially explicit model of sapling growth in a tropical forest: does the identity of neighbours matter? Journal of Ecology. 92(2):348-360.
Vincent, G., and D. Harja. 2007. Exploring Ecological Significance of Tree Crown Plasticity through Three-dimensional Modelling. Annals of Botany. 101(8):1221-1231.
Vourtsas, G., I. Evangelidou, and G. Intzidou. 2014. Study of development – protection – management of the forest ecosystem of Kastaneris – Livadion. Kilkis Forest Service.
Watkinson, A., W. Lonsdale, and L. Firbank. 1983. A neighborhood approach to self-thinning. Oecologia. 56(2-3):381-384.
Weiner, J., and S. Thomas. 1992. Competition and Allometry in Three Species of Annual Plants. Ecology. 73(2):648-656.
Weiskittel, A., D. Hann, Jr. Kershaw, and J. Vanclay. 2011. Forest Growth and Yield Modeling. Wiley & Sons, Chichester, UK.
Zerva, A., G. Halyvopoulos, and K. Radolgou. 2008. Fine root biomass in a beech (Fagus sylvatica L.) stand on Paiko Mountain, NW Greece. Plant Biosystems. 142(2):381-385.



