Schlüsselbegriffe: Mischwaldbewirtschaftung, Umbau, Wiederherstellung, Degradation, Arten Diversität
Abstract
Mixed-species forest management has been receiving increasing attention due to the diverse ecological and economic benefits. Afforestation with multiple tree species, promoting mixed species through natural regeneration and the transformation of planted monocultures to mixed stands are the main focus of mixed-species forest management. The optimal species configuration, i.e., an appropriate spatial arrangement of young and adult trees of different species, is of great importance to such transformations, as well as to the restoration of degraded secondary natural forests. Species association refers to a recurring group of co-occurring species. Species pairs with positive association share the same habitat requirement and thus coexist well. The measurement of species association facilitates a comprehensive understanding of community structure, function, formation, maintenance and succession in the forest. In turn, the association of the tree species in a given area can be a measure for species configuration. Little is known about this species association in natural forests in the western Hunan Province, China. In this study, the species association between dominant tree species (including sapling and adult trees) was investigated in an old-growth forest. The overall species association was first determined using a variance ratio (VR) test and the pairwise species association was then determined using a chi-square (χ2) test, the association coefficient (AC), the Dice index (DI), and Spearman’s rank correlation coefficient (SRCC). The VR test showed a significant overall positive species association, indicating a stable structure in the old-growth forest. We also determined the pairwise intra- and interspecific association between saplings and adult trees. The light requirements of the tree species were considered and the optimal species configuration was determined, important for the establishment of mixed-species plantations, the transformation of monocultures, and enrichment plantings in degraded secondary natural forests. Additionally, based on the findings of this study we provided suggestions for suitable silvicultural practices.
Zusammenfassung
Die Waldbewirtschaftung von Mischbeständen hat in der Vergangenheit aufgrund der vielfältigen ökologischen und wirtschaftlichen Vorteile verstärkte Aufmerksamkeit erhalten. Die Aufforstung mit mehreren Baumarten und der Umbau von Reinbeständen sind wesentliche Aspekte der Bewirtschaftung in Mischwäldern. Die optimale Artenkonfiguration, das bedeutet eine geeignete räumliche Anordnung von jungen Bäumen und erwachsenen Bäumen verschiedener Arten, ist für solche waldbaulichen Bestrebungen sowie für die Wiederherstellung degradierter sekundärer Naturwälder von großer Bedeutung. Artenassoziation bezieht sich auf eine wiederkehrende Gruppe von gleichzeitig vorkommenden Arten. Artenpaare mit positiver Assoziation teilen den gleichen Lebensraumbedarf und ko-existieren daher gut. Die Messung dieser Artenassoziation ermöglicht ein umfassendes Verständnis der Struktur, Funktion, Entstehung, Erhaltung und Dynamik von Waldgesellschaften. Somit ist die Artenassoziation der Baumarten in einem bestimmten Gebiet ein Werkzeug für die Beurteilung der Artenkonfiguration. Über die räumliche Zuordnung der Baumarten in natürlichen Wäldern in der westlichen Provinz Hunan in China ist jedoch wenig bekannt. In dieser Studie wurde die Artenassoziation zwischen dominanten Baumarten (Verjüngung und ausgewachsenen Bäumen) in einem Urwald untersucht. Die Gesamtartenassoziation wurde zuerst mittels eines Varianzverhältnis (VR)-Tests bestimmt, und die paarweise Artenassoziation wurde dann mit einem Chi-Quadrat-Tests (χ2), Assoziationskoeffizienten (AC), Dice-Index (DI) und Spearman-Rangs Korrelationskoeffizient (SRCC) bestimmt. Der VR-Test zeigte eine insgesamt signifikante positive Artenassoziation, was auf eine stabile Struktur und Artenzusammensetzung im Urwald hindeutet. Wir haben auch die paarweise intra- und interspezifische Assoziation zwischen Verjüngung und ausgewachsenen Bäumen bestimmt. Der Lichtbedarf der Baumarten wurde berücksichtigt und die optimale Artenanordnung für die Etablierung von Mischbeständen, für den Umbau von Reinbeständen und für die Ergänzungspflanzungen in degradierten sekundären Naturwäldern ermittelt. Außerdem wurden basierend auf den Ergebnissen dieser Studie Vorschläge für geeignete waldbauliche Praktiken gemacht.
1. Introduction
Natural forests are an important component of terrestrial ecosystems because they provide many ecological functions such as soil and water conservation, biodiversity, water and air purification, wildlife habitat, and wood and non-wood forest products. (Bruijnzeel 2004; De Groot et al. 2002; Hansen et al. 1995; Pimentel et al. 1992). The western Hunan Province China is located in the mid-subtropical zone and has diverse species richness and a unique climate (Li et al. 2008; Xiong et al. 2007). However, in the past, forest management was focused on monocultures with the objective of timber production, whereas management and conservation of natural forests were not considered. Moreover, the natural forests in western Hunan Province have historically suffered from long-term and extensive anthropogenic disturbances, e.g., unsustainable exploitation and illegal logging, resulting in poor forest quality (Li 2004; Wang et al. 2001; Yang 2001). Therefore, natural forests in this area are mainly degraded secondary forests that do not meet the increasing demand for ecological and economic benefits of forests. For instance, according to the 9th Chinese National Forest Inventory (NFI), the average stocking volume in degraded secondary forests is only 45.32 m3 per ha in western Hunan Province. In 2017, China imposed a total ban on commercial logging in natural forests (Hou et al. 2019). Under these conditions, forest managers have to deal with considerable challenges when planning science-based and sustainable management of degraded natural forests with the goal to quickly improve forest quality. It has been demonstrated that enrichment planting with native tree species is a promising approach to restoring degraded secondary forests (Mangueira et al. 2019; Tigabu et al. 2010; Ådjers et al. 1995; Ramos 1992). Many authors have investigated optimal enrichment planting and related silvicultural practices (Mangueira et al. 2019; Tigabu et al. 2010). For instance, Mangueira et al. (2019) investigated enrichment planting methods to restore tropical forest fragments in the Brazilian Atlantic Forest and concluded that a combination of planting methods at the species level was likely to increase the restoration success.
Western Hunan Province holds 5.02 million ha of plantations in, comprising 47.67 % of the total forest cover of the province. Chinese fir (Cunninghamia lanceolata) and Masson pine (Pinus massoniana) are the two dominant tree species used in plantations in this area. Plantations have been criticized because they are often ecologically unstable forests, provide a low level of ecological services, and are highly susceptible to disturbance (Wang 2018; Felton et al. 2010; Richards et al. 2010) . For instance, in February 2008, about 18 million ha of plantations in the subtropical regions of China were affected by heavy sleet and ice, causing economic losses of 57.3 billion Chinese Yuan (around 9.3 billion US Dollar) (Meng et al. 2014). Many authors have documented that mixed-species forests have many advantages over monoculture plantations in terms of providing ecological services such as carbon sequestration, biodiversity conservation, and wildlife habitat protection (Felton et al. 2016; O'Hara 2014; Wang et al. 2008; O'Hara 1998). In addition, Huang et al. (2018) reported that there was a significant positive correlation between tree species diversity and forest productivity and suggested that the management of mixed-species forests was a promising approach for maintaining biological conservation and mitigating climate change. Mixed-species forests can be established by afforestation with multiple tree species. A large number of successful cases of mixed-species plantations have been documented in China (He et al. 2013; Wang et al. 2013; Jiang et al. 2010; Wang et al. 2008). Mixed-species forests can also be created by transforming monocultures by thinning and enrichment planting of various native tree species (Wilson et al. 2018; Meng et al. 2014). In China, a large number of studies have been conducted on the transformations of monocultures using various tree species, e.g., Cunninghamia lanceolata (Sun et al. 2015; Meng et al. 2014), Pinus massoniana (Meng et al. 2014; Luo et al. 2013), Pinus tabuliformis (Ning et al. 2009), and Pinus yunnanensis (Zhang et al. 2011; Cai et al. 2006).
The selection and spatial configuration of mixed tree species is a key prerequisite for establishing mixed-species plantations, transforming monocultures, and conducting enrichment planting to restore degraded secondary forests. Species association is defined as a recurring group of co-occurring species (Legendre 1978). Species association indicates the spatial distribution relationship and functional dependency between different species and is a static description of the relationship of plant community species over a certain period (Jian et al. 2009; Lin et al. 2005; Peng et al. 1999; Greig-Smith 1983; Cole 1949). Species association can be categorized as overall association and pairwise association (Su et al. 2015; Ludwig et al. 1988). An overall positive association indicates that the population of dominant species exhibits a stable phase and appears to exist in a mutually beneficial relationship (Guo et al., 2017; Su et al. 2015). Species pairs with significant positive associations tend to have similar ecological characteristics under the same site conditions and there is a high probability that they are closely associated and co-exist well (Zhao et al. 2012; Hao et al. 2007). Therefore, species association may provide scientific evidence for identifying the optimal spatial configuration of mixed tree species. In fact, species associations have been extensively examined in the support of forest management. Hao et al. (2007) investigated the intra-species spatial relationships between adult and juvenile trees of different species and determined the optimal species configuration for enrichment plantings. Zhao et al. (2012) investigated species associations among saplings and between saplings and adult trees and provided recommendations for the selection and configuration of tree species for enrichment planting in spruce-fir forests in the Changbai Mountains. The species association of different tree species in western Hunan Province, has not been studied so far.
An old-growth forest is defined as a forest that has attained great age without significant disturbance and exhibits unique ecological features; it is commonly classified as a climax community (White 1994). Since historically old-growth forests did not experience any management, they provide unique opportunities to conduct studies that cannot be performed in managed forests.
In this study, we first investigated intra- and interspecies spatial associations between saplings and adult trees in an old-growth forest in western Hunan Province. Secondly, based on the species association results, we propose an optimal species configuration between the saplings and adult trees to support the establishment of mixed-species plantations, the transformation of monocultures, and the enrichment planting in degraded secondary natural forests.
2. Materials and Methods
2.1 Study area
The study area is located at the Huitong Experimental Station of Forestry Ecology of the Chinese Academy of Sciences (26°50’N, 109°36’E) (Fig. 1).
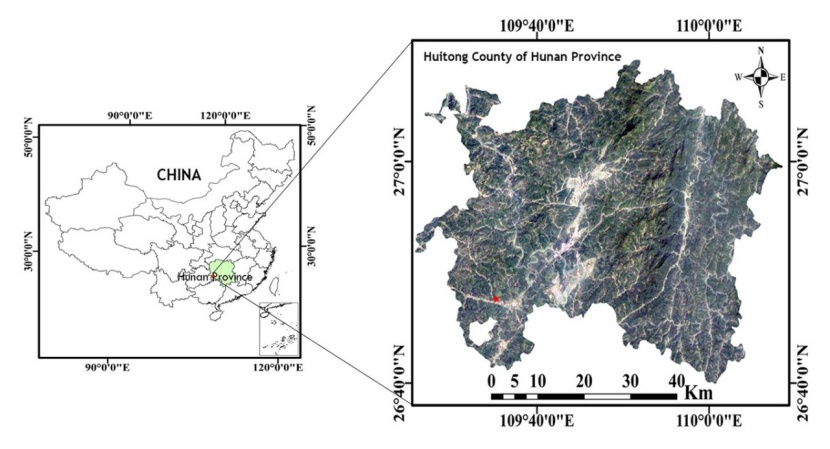
Figure 1: The location of our study site at the Huitong Experimental Station of Forestry Ecology of the Chinese Academy of Sciences. The red star represents the center of the Huitong Experimental Station. / Abbildung 1: Standort unserer Versuchsflächen an der Huitong Versuchsstation für Forstökologie der Chinesischen Akademie der Wissenschaften in der westlichen Provinz Hunan. Der rote Stern zeigt die Lage der Huitong Versuchsstation.
The region has a subtropical monsoon humid climate with an annual precipitation of 1200-1400 mm, a mean annual air temperature of 23.6°C, and annual sunshine hours ranging from 1,300 to 1,800 hours. The altitude range of the area is 300-415 m. Red-yellow soil is the most common soil type in this region, which has developed from the slate and shale (Deng et al. 2007; Wang et al. 2007). The natural zonal vegetation in this area is subtropical evergreen broad-leaved forest dominated by Castanopsis spp. and Lithocarpus spp. The area is rich in tree species and most are endemic species. The tree species include Castanopsis fargesii, Cyclobalanopsis glauca, Machilus pauhoi, Engelhardia roxburghiana, Meliosma rigida, Litsea mollis, and Vernicia montana. The dominant shrub species are Maesa japonica, Camellia japonica, and Eurya japonica. The dominant herb species are Woodwardia japonica and Smilax microphylla.
2.2 Description of old-growth forest and plot establishment
The studied old-growth evergreen broad-leaved forest is located in an extremely remote area, where there has been largely no anthropogenic disturbance historically. Therefore, this old-growth forest represents the local climax vegetation community. In 1997, a square plot with an area of 2,500 m2 was established. Then, the plot was divided into twenty-five 10 m × 10 m sub-plots. In each subplot, we recorded the diameter at breast height at 1.3 m (DBH), tree height (H), and location of all trees greater than 1 cm in DBH. For all trees we identified the species. The plots were re-measured every year. Based on height and diameter, the trees were divided into saplings (DBH < 5.0 cm and H ≥ 1.3 m) and adult trees (DBH ≥ 5.0 cm and H ≥ 1.3 m) (Zhang et al. 2012). The association analysis is based on the latest measurement data in 2018. A descriptive statistics of this old-growth forest stand was summarized in Table 1.
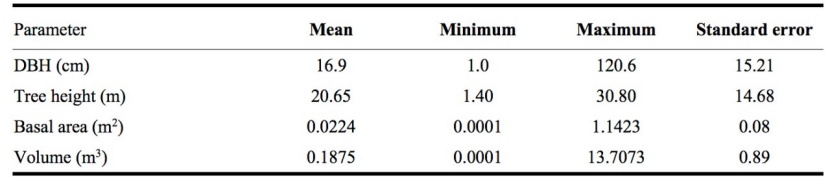
Table 1: Summary of stand characteristics of the old-growth forest with individual.
Additionally, the diameter distribution of the old-growth forest was also provided in Fig 2.
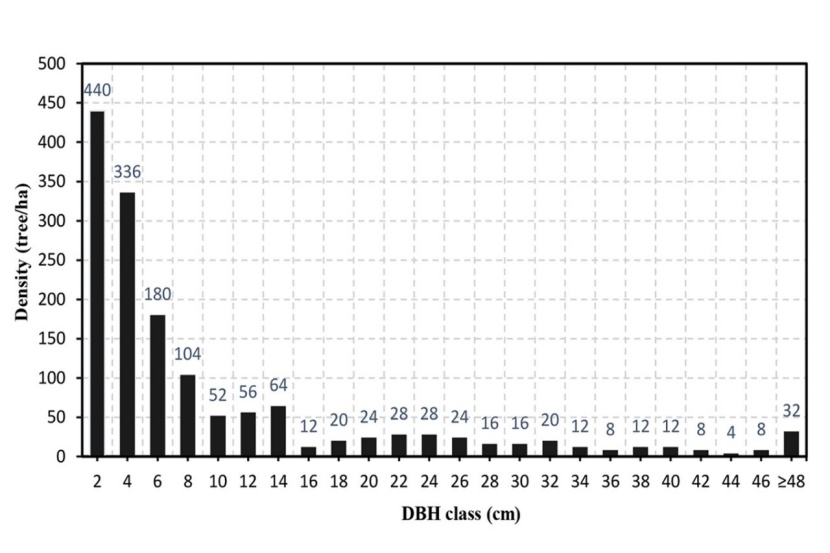
Figure 2: Diameter distribution of the old-growth evergreen broad-leaved forest. / Abbildung 2: Durchmesserverteilung des immergrünen Laub-Urwaldes.
2.3 Method
2.3.1 Importance value
The importance value (IV) is a comprehensive quantitative indicator used to characterize the status and role of each species in the community (Legendre 1978; Isango et al. 2007; Erman 1971). The larger the IV of a tree species, the more dominant the species is in the plot. The IV was calculated as follows:

2.3.2 Overall species association analysis
We first investigated the overall association between all possible species pairs of saplings and adult trees in the old-growth forest; if a significant overall association was observed, then the pairwise species association was determined. In this present study, we used the variance ratio (VR) test (Gu et al. 2017; Ludwig et al. 1988; Schluter 1984), which is based on species presence or absence, to examine the overall association. The VR is calculated as follows:
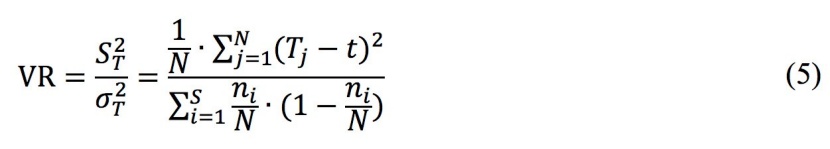
where S is the total number of species, N represents the total number of quadrats, Tj is the number of species in the jth quadrats, ni denotes the number of quadrats in which the ith species occurred, and t is the average number of species in the quadrats.
Under the null hypothesis of independence, the expected value of VR is 1. When VR > 1, the species exhibits a positive association and when VR < 1, a negative association is observed (Gu et al. 2017; Ludwig et al. 1988) . Additionally, we computed the statistic W = N×VR to test whether the deviations of the VR from 1 were significant. If there is no significant species association, there is a 90 % probability that W lies between the limits given by the chi-square (χ2) distribution: χ20.05 ≤ W ≤ χ20.95 (Ludwig et al. 1988).
2.3.3 Pairwise species association analysis
The VR test only shows whether there is an overall species association in a forest stand but does not reveal pairwise intra- and interspecies spatial associations. Therefore, in this study, once the overall species association was identified, the χ2 statistics, the association coefficient (AC), the Dice index (DI) and Spearman’s rank correlation coefficient (SRCC) were used to determine the species spatial association between all possible species pairs of saplings and adult trees.
2.3.3.1 The chi-square (χ2) test
The χ2 statistic was based on a 2×2 contingency table. The χ2 test statistic will be biased if any cell in the 2×2 contingency table has an expected frequency of less than 1 or if more than two of the cells have expected frequencies of less than 5 (Zar 1974). Therefore, Yate’s correction formula was employed to correct the χ2 statistic to ensure closer approximation to the theoretical, continuous χ2 distribution. The corrected χ2 statistic is calculated as follows:
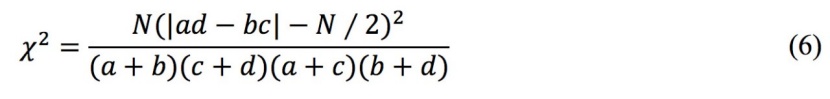
where N is the total number of quadrats, a is the number of quadrats in which both A and B (species A and B) are present, b denotes the number of quadrats in which only A appears, c represents the number of quadrats in which only B appears, and d is the number of quadrats without A or B. When χ2 < 3.841, the species are independent of each other and no species association is detected; when 3.841 ≤ χ2 ≤ 6.635, there is a significant association between the species; when χ2 > 6.635, there is a highly significant association between the species. When ad-bc > 0, the association is positive and when ad-bc < 0, the association is negative.
2.3.3.2 The association coefficient (AC) and Dice index (DI)
The (χ2) test only provides a qualitative evaluation of the pairwise species association but does not quantitatively describe the degree of the association. Furthermore, there might be a species association between two species, even though the χ2 test showed no significance (Xue 2016). Many authors suggested that the χ2 test should be used in conjunction with the AC, which provides a quantitative measure of the degree of species association (Gu et al. 2017; Ye et al. 2017; Su et al. 2015). The AC is calculated as follows:
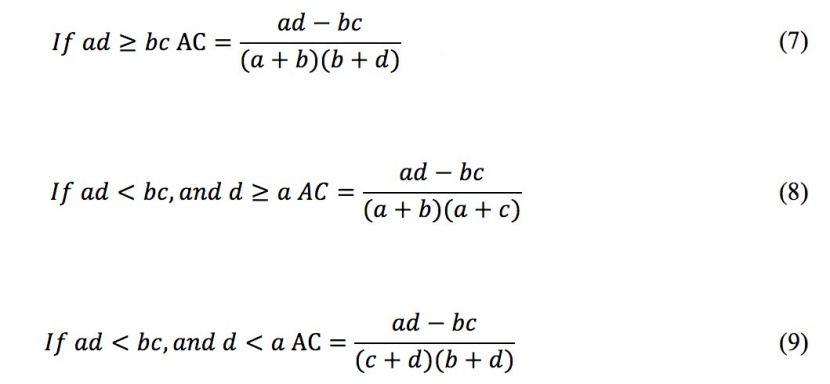
The AC range is [-1, 1]; if AC = 1, the positive species association is strongest between the species pairs and AC = -1 denotes that the species pair has the strongest negative association, whereas AC = 0 indicates that the species pair is completely independent.
Since the AC is highly influenced by the d (the number of quadrats without two species), bias can occur (Liu et al. 2017). Therefore, we also included the DI, which removes the bias caused by the AC, to quantitatively represent the species association (Liu et al. 2017; Ludwig et al. 1988). The DI is calculated as follows:

The DI range is [0,1]. When DI = 0, it indicates the complete independence of the species pairs and the species do not appear in the same quadrat; the closer the DI is to 1, the higher is the probability that the species pairs appear in the same quadrat. We employ the R package “spaa” and “plyr” to derive chi-square (χ2) statistics, AC and DI.
2.3.3.3 Spearman’s rank correlation coefficient (SRCC)
Although the AC and DI quantitatively describe the degree of species association, they are constructed using binary data; therefore, inevitably some information is lost, e.g., species abundance (Ludwig et al. 1988). Ludwig et al. (1988) stated that if species abundance data are available, the species covariation should be determined. Although Hurlbert (1969) argued that it was important to make a sharp distinction between species association and species covariation, unfortunately, these two terms are often inappropriately interchanged. Since species covariation can provide information on how two species co-vary, in this study, we used the updated version of species association. Ludwig et al. (1988) also stated that the term correlation was general but in an ecological context, it should be reserved for how two species co-vary. In the present study, we used the SRCC to investigate the species covariation. The SRCC, which uses the rank of the data, is a nonparametric technique for evaluating the degree of linear association or correlation between independent variables. Its advantages over other correlation coefficients include insensitivity to the distribution of the population and outliers and tolerance of a very small sample size (Gautheir 2001). The SRCC is calculated as follows:

where rs (i, j) is the Spearman’s rank correlation coefficient, N is the total number of quadrats, dk = (xik-xjk), xik is the rank of species i in quadrat k, and xjk is the rank of species j in quadrat k. We calculated the SRCC using the R Package “plyr” and “corrplot”.
3 Results
3.1 Species composition
There were a total of 36 tree species with an average DBH of 16.90 cm and an average height of 20.65m in the natural old-growth forest. The ranking of the importance values (IVs) of the tree species is shown in Table 2. Castanopsis fargesii was the most dominant tree species in the old-growth forest and its IV was 2.44 times that of the second dominant tree species, i.e., Machilus pauhoi. Because there were fewer individuals of tree species with a small IV, we only investigated the species association for the 10 tree species with the highest IVs. These 10 tree species represented 81.26 %, 84.69 %, and 78.74 % of the relative abundance, relative dominance, and IVs, respectively, indicating their dominance in the old-growth forest. In addition, the light requirements of the tree species in the old-growth forest are also listed in Table 2.
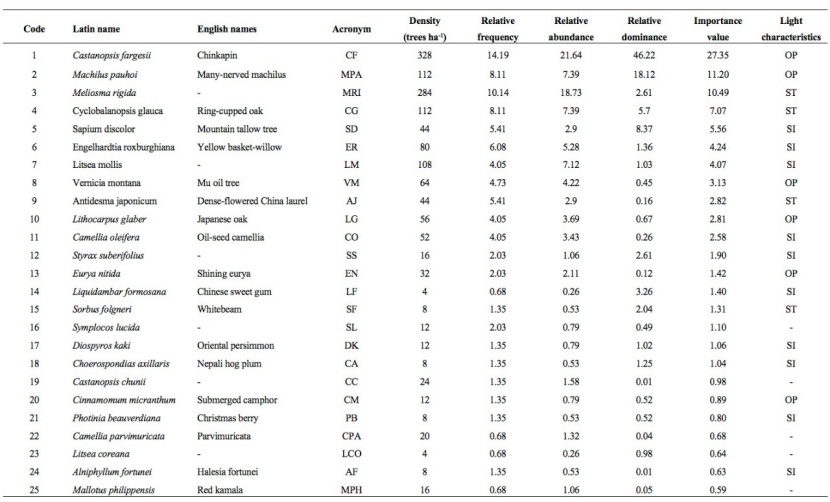
Table 2: The importance values and light requirements of the tree species in the old-growth forest.
3.2 Overall species association in the old-growth forest
The VR is 2.033, indicating an overall net positive species association in the old-growth forest. Moreover, the W statistics with the value of 50.832 fell outside the confidence interval. Therefore, it appears that the forest is in a relatively stable state in terms of its structure and species composition.
3.3 Pairwise species association in the old-growth forest
3.3.1 Test of pairwise species association
The pairwise species association was investigated using the χ2 test and the results are shown in Fig. 3. The results showed that 80 pairs exhibited a positive association and 91 pairs exhibited a negative association, accounting for 46.78 % and 53.22 % of the total number of pairs, respectively. Among the 171 tree species pairs, 3 pairs exhibited highly significant positive associations, which were the S. discolor (U) - E. roxburghiana (U), L. mollis (S) - L. mollis (U), V. montana (S) - L. mollis (U), and 3 pairs exhibited significant positive associations, which were M. rigida (S) - S. discolor (U), V. montana (S) - V. montana (U), M. pauhoi (S) - E. roxburghiana (U). We used the abbreviations S and U for saplings and adult trees. In addition, the χ2 test also indicated that 2 pairs showed significant negative associations, which were M. rigida (S) - L. mollis (S) and M. rigida (S) - L. mollis (U).
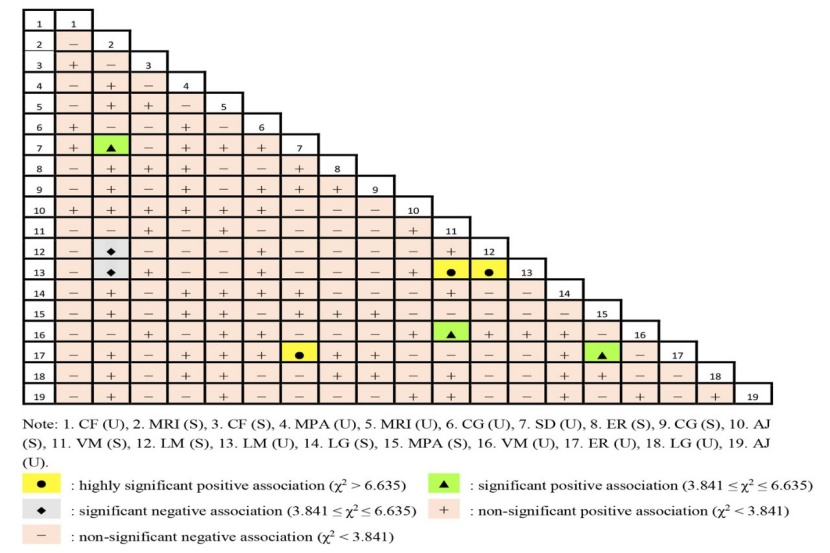
Figure 3: Correlation matrix of the χ2 test results of the dominant species in the studied old-growth forest. / Abbildung 3: Korrelationsmatrix der χ2-Testergebnisse der vorherrschenden Arten im untersuchten Urwald.
3.3.2 Measure of pairwise species association using the AC and DI
The correlation matrix of the AC values is shown in Fig. 4. We observed that among the 171 tree species pairs, 16 pairs exhibited strong positive associations (AC ≥ 0.6) and 43 pairs exhibited weak positive associations (0.2 ≤ AC < 0.6), accounting for 9.4 % and 25.1 % of the total species pairs, respectively. In contrast, 28 pairs showed weak negative associations (-0.6 < AC ≤ -0.2) and 46 pairs showed strong negative associations (AC ≤ -0.6), accounting for 16.4 % and 26.9 % of the total species pairs, respectively. In addition, there were 38 pairs that had no associations (-0.2 < AC < 0.2).
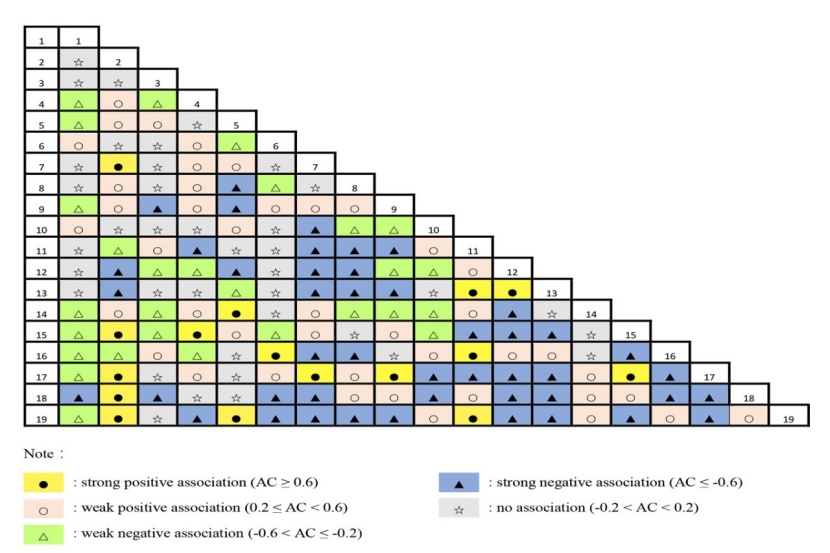
Figure 4: Correlation matrix of the AC of the dominant species in the studied old-growth forest. / Abbildung 4: Korrelationsmatrix der AC der vorherrschenden Arten im untersuchten Urwald.
The DI was also calculated to obtain a quantitative measure of the pairwise species associations; the results are shown in Fig. 5. Amongst the 171 species pairs, 10 pairs (5.9 %) exhibited a very strong associations (DI ≥ 0.6), which were C. fargesii (S) - C. fargesii (U), M. rigida (S) - M. pauhoi (U), M. rigida (S) - M. rigida (U), M. rigida (S) - S. discolor (U), C. fargesii (S) - M. rigida (U), V. montana (S) - L. mollis (U), V. montana (S) - V. montana (U), L. mollis (S) - L. mollis (U), M. pauhoi (S) - E. roxburghiana (U), and S. discolor (U) - E. roxburghiana (U). There were 37 pairs with strong associations (0.4 ≤ DI < 0.6) and 55 pairs with weak associations (0.2 ≤ DI < 0.4), accounting for 21.6 % and 32.2 % of the total tree species pairs, respectively. In addition, 69 pairs (DI < 0.2) had no association, which accounted for 40.3 % of the total species pair.
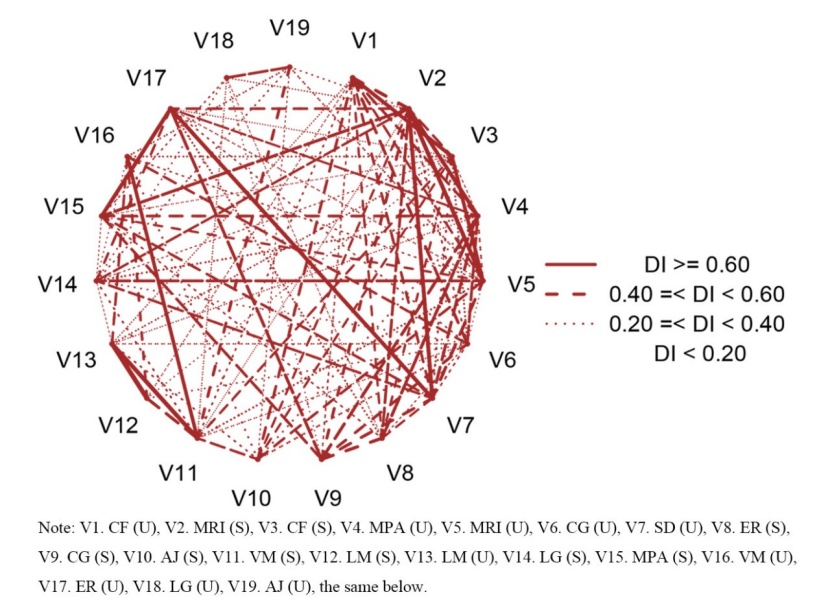
Figure 5: Network of the DI of the dominant species in the studied old-growth forest. / Abbildung 5: Netzwerk der DI der vorherrschenden Arten im untersuchten Urwald.
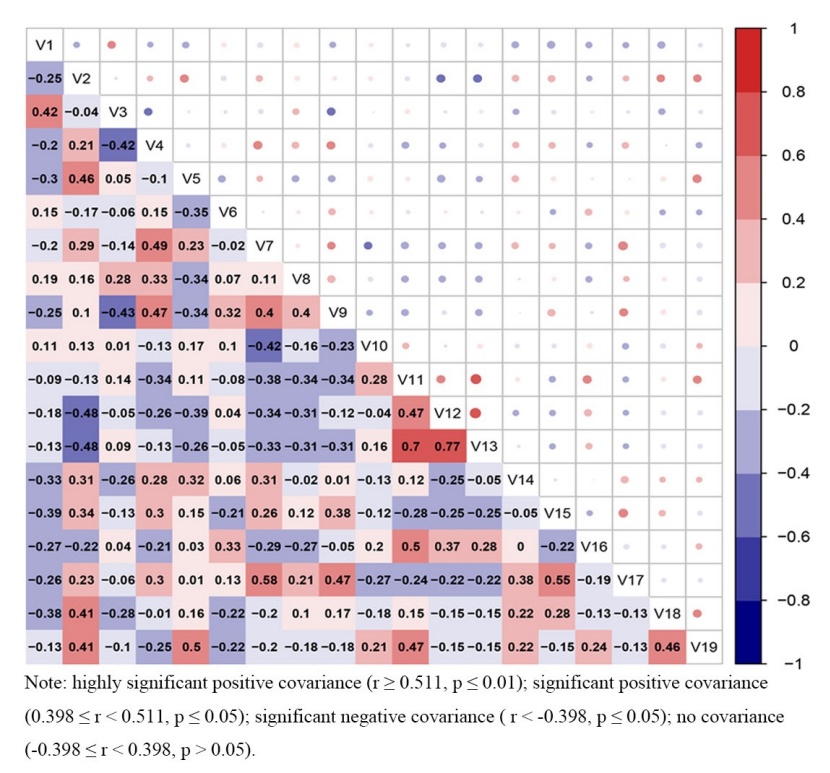
Figure 6: Correlation matrix of the SRCC of the dominant species in the studied old-growth forest. / Abbildung 6: Korrelationsmatrix des SRCC der vorherrschenden Arten im untersuchten Urwald.
3.3.3 Measure of pairwise species covariation using the SRCC
The species covariation was investigated using the SRCC and the results are listed in Fig. 6. Among the 171 species pairs, 77 pairs exhibited a positive covariance, 93 pairs exhibited a negative covariance and 1 pair showed no covariance, accounting for 45.0 %, 54.4 %, and 0.6 % of the total pairs, respectively. Four pairs exhibited highly significant positive covariance (rs ≥ 0.511, p ≤ 0.01), i.e., V. montana (S) - L. mollis (U), L. mollis (S) - L. mollis (U), M. pauhoi (S) - E. roxburghiana (U) and S. discolor (U) - E. roxburghiana (U), whereas 14 pairs exhibited significant positive covariance (0.398 ≤ rs < 0.511, p ≤ 0.05), i.e., C. glauca (S) - E. roxburghiana (S), L. mollis (S) - V. montana (S), C. fargesii (S) - C. fargesii (U), M. rigida (S) - M. rigida (U), M. rigida (S) - L. glaber (U), M. rigida (S) - A. japonicum (U), C. glauca (S) - M. pauhoi (U), glauca (S) - S. discolor (U), glauca (S) - E. roxburghiana (U), V. montana (S) - A. japonicum (U), V. montana (S) - V. montana (U), M. pauhoi (U) - S. discolor (U), A. japonicum (U) - M. rigida (U), and A. japonicum (U) - L. glaber (U). Additionally, 5 pairs showed a significant negative covariance (rs < -0.398, p ≤ 0.05), i.e., M. rigida (S) - L. mollis (S), C. fargesii (S) - C. glauca (S), M. rigida (S) - L. mollis (U), C. fargesii (S) - M. pauhoi (U), and A. japonicum (S) - S. discolor (U).
4. Discussion
4.1 Final determination of species association
The overall species association describes the static relationship among all species in the community and the overall significant positive association suggests that the community is in a relatively stable state (Gu et al. 2017; Su et al. 2015). In this study, The VR was 2.033, which was larger than 1, i.e., the expected value of the VR under the null hypothesis of species independence. Moreover, the W statistics further confirmed the significant overall positive species association, indicating the stable structure and species composition in the old-growth forest. Indeed, the relative stability of old-growth forests was extensively documented by many authors (Kimmins 2003; Spies 1996; Thomas et al. 1988).
The χ2 test showed that there was little difference between the number of pairs with positive associations and negative associations and in most of the pairs, the association was not significant. It may be that the species competition in the old-growth forest was within the environmental capacity and hence the forest community was in a stable state (Hu et al. 2013). Additionally, the fact that there were only a few pairs with statistically significant species associations may be attributed to the χ2 test itself. For instance, Xu (2016) argued that although the χ2 test can determine whether the species association is significant or not, it cannot measure the degree of association. Even if a species association may be present, the χ2 test did not detect it. Unlike the χ2 test, the AC and DI do provide quantitative measures of the relative degree of the species association but there is a lack of a unified standard for defining the levels of strength and weakness, resulting in different conclusions for different standards (Guo et al. 1997). For instance, Su et al. (2015) defined the following 5 levels of AC, i.e., strong positive association (0.6 < AC ≤ 1.0), weak positive association (0.2 < AC ≤ 0.6), no association (-0.2 < AC ≤ 0.2), weak negative association (-0.6 < AC ≤ -0.2), and strong negative association (-1.0 ≤ AC ≤ - 0.6). In comparison, 4 levels of AC, i.e., 0.5 < AC ≤ 1.0, 0.1 ≤ AC ≤ 0.5, -0.1 ≤ AC ≤ -0.5, and -1.0 ≤ AC < -0.5 were proposed by Chai et al. (2016) and Zhang et al. (2015). Although the AC and DI result in a better performance than the χ2 test in describing the species association, it is noteworthy that, similar to the χ2 test, they are based on binary data, which inevitably leads to a loss of information.
In the SRCC, continuous data are used to evaluate the degree of linear association or correlation between independent variables (Ludwig et al. 1988). Furthermore, it is worth noting that the SRCC is a nonparametric technique that is unaffected by the distribution of the population (Gautheir 2001; Ludwig et al. 1988). Therefore, the SRCC is well suited to be used in conjunction with the χ2 test for determining species association. In this study, significant SRCC values were obtained for species pairs that had significant χ2 values. In addition, the number of significant species pairs was higher for the SRCC test than the χ2 test. The reason may be attributed to the fact that the χ2 test is based on binary data, whereas the SRCC is based on continuous data; this results in a higher sensitivity of the SRCC to determine the co-occurrence of two species (Li et al. 2001). Many authors suggested that the combination of the χ2 test and the SRCC was necessary to provide reliable results (Zhou et al. 2002; Dezfuli et al. 2001; Li et al. 2001; Mundry 1998).
Consequently, in this study, the final species association was based primarily on the χ2 test and the SRCC and using AC and DI as supporting information. Our results showed that for the pairs that were significant in the χ2 and the SRCC tests, the AC and DI values were also significant. The pairs that exhibited highly significant positive and negative associations in the χ2 and the SRCC tests were listed in Table 3. Overall, we are very confident with the results of the species association of these pairs because they passed all four tests. In turn, they will be given first priority for the species configuration.
The species pairs that exhibited non-significant associations according to the χ2 test but significant associations based the SRCC, AC, and DI were listed in Table 3. Li et al. (2001) reported that, even though some species pairs were not significant in the χ2 test, it did not absolutely mean that they are not associated. These tree species pairs will be given second priority for the species configuration.
The pairs that showed significant associations based only on the SRCC were listed in Table 3. All these pairs except C. fargesii (S) - M. pauhoi (U) had significant positive associations. These pairs will be given third priority for the species configuration. Additionally, the species pair M. rigida (S) - S. discolor (U) had a significant positive association in the χ2 test, AC, and DI but the SRCC was not significant. The species pair M. rigida (S) - M. pauhoi (U) had relatively high AC and DI values but the association was not statistically significant in the χ2 and SRCC tests. These tree species pairs will be given fourth priority for the species configuration (Table 3).

Table 3: The classification of species association pairs for the species arrangement.
4.2 Possible reasons for the observed species associations
The species pairs with significant positive associations tend to have similar ecological characteristics under the same site conditions and there is a high probability that they are closely associated and coexist well (Zhao et al. 2012; Hao et al. 2007). This was confirmed in our study and examples include the drought and barren-tolerant species pair C. glauca (S) - E. roxburghiana (S), the heliophilous, fertile-, and acid-tolerant species pair V. montana (S) - L. mollis (S), the shade-tolerant and hygrophilous species pair M. rigida (U) – A. japonicum (U), and the heliophilous and hygrophilous species pair S. discolor (U) - E. roxburghiana (U).
We observed that the saplings of C. fargesii (S), M. rigida (S), V. montana (S), and L. mollis (S) had significant positive correlations with the adult of the same species. Similar results were also reported by many other studies. For instance, Hao et al. (2007) reported a positive association between the saplings and adult trees of Quercus wutaishanica. Du et al. (2013b) observed a positive association between Pinus koraiensis in the lower forest layer and the dominant layer. Many authors attributed this to the limited seed dispersal ability and the shade tolerance of the saplings (Hao et al. 2007; Hou et al. 2004).
In addition to the positive intraspecific association between saplings and adult trees, we also detected the significant positive interspecific associations between saplings and adult trees; examples include M. pauhoi (S) - E. roxburghiana (U), C. glauca (S) - S. discolor (U), and V. montana (S) - L. mollis (U). This positive interspecific association between saplings and adult trees might be explained by the niche differentiation in terms of resource utilization for these species pairs (Yuan et al. 2018; Sushma 2006). Additionally, it is worth noting that the saplings of M. rigida (S) and C. glauca (S) had significant positive associations with the adult trees of many other tree species. Hao et al. (2007) attributed this phenomenon to the relatively broad niche of saplings of many species, which allows them to grow well under the canopy of many other trees species.
In addition to the positive association, the following negative associations amongst pairs were observed, i.e., C. fargesii (S) - C. glauca (S), C. fargesii (S) - M. pauhoi (U), A. japonicum (S) - S. discolor (U), M. rigida (S) - L. mollis (S), and M. rigida (S) - L. mollis (U). These negative associations reflected the different habitat requirements of the two tree species in the pair and their interspecific competition in case of resource scarcity. Similar findings have been reported in many studies. For instance, Deng et al. (2015) reported highly significant negative associations between Ulmus laciniata and Tilia amurensis and attributed the negative association to the different characteristics of the two species, i.e., U. laciniata is adapted to drought and saline-alkali soil, whereas T. amurensis is not. However, different habitat requirements did not totally mean that negative association existed and it might also lead to positive facilitation effects. A specific example is the niche complementary theory, which states that plant species or functional groups occupy functionally distinct niches in an ecosystem and use resources in a complementary way (Kahmen et al. 2006). For instance, Amoroso (2006) and Manson et al. (2006) documented that in mixed-species tree plantations canopy stratification between shade-tolerant and intolerant species reduces light competition and increases light interception.
4.3 Implications for silvicultural practices
Information on species associations provides a scientific basis for species configuration in plantings and is of great importance for forest management (Zhao et al. 2012; Hao et al. 2007).In addition, the light requirements of tree species also have to be considered in the species configuration and forest management decision making (Wang 2018; Klopčič et al. 2015; Van Couwenberghe et al. 2013). In this study, we categorized the tree species into shade-intolerant species, shade-tolerant species, and opportunistic species according to their light requirements (Wang 2018). Among the 10 dominant tree species in the old-growth forest, the shade-tolerant species included M. rigida, C. glauca, and A. japonicum; the opportunistic species included C. fargesii, M. pauhoi, V. Montana, and L. glaber; the shade-intolerant species included S. discolor, E. roxburghiana, and L. mollis. Based on the species associations and their light requirements, we provide the following suggestions for species configuration for establishing mixed-species plantations, transforming monocultures, and conducting enrichment planting to restore degraded secondary forests. Additionally, silvicultural recommendations are also provided.
The objective of afforestation is to rapidly establish a forest stand. Therefore, in this case, we recommend shade-intolerant species, which include S. discolor, E. roxburghiana, L. mollis, and L. formosana. Additionally, some opportunistic species can also be considered, such as V. montana, C. fargesii, and L. glaber since they have high adaptability and robustness due to their light requirements and ability to establish on bare land (Wang 2018). The study results indicated that the pairs of C. glauca (S) - E. roxburghiana (S) and S. discolor (U) - E. roxburghiana (U) had positive associations. Even though C. glauca is a shade-tolerant species, some studies have documented that it was suitable for afforestation due to its drought tolerance, ability to establish on bare soil, and high adaptability (Yuan et al. 2012). Therefore, we propose to establish mixed-species plantations of C. glauca and E. roxburghiana, as well as S. discolor and E. roxburghiana.
In addition, in harsh site conditions, we strongly suggest to first establish a P. massoniana plantation to increase the tree survival rate because this species is fast-growing and drought- and barren-tolerant (Du et al. 2018; Liu et al. 2006). When a closed canopy has been established, the pure P. massoniana plantation can be diversified by using an enrichment planting of positively associated tree species pairs such as L. mollis (S) - V. montana (S) and C. glauca (S) - E. roxburghiana (S). These saplings grow rapidly, have high adaptability, and provide forest species diversity. The dynamics of the enrichment plantings should be carefully monitored and forest gaps should be created in a timely manner to facilitate good forest growth.
The transformations of monocultures are an important method for creating mixed-species forests. The objective of the transformation is to convert monocultures into an uneven-aged mixed-species heterogeneous forest to achieve continuous crown cover (Wang 2018; Meng et al. 2014; Adams et al. 2011; O’Hara 2001). The most important task of plantation transformations is to fast establish regeneration layers (Wang 2018). The shelterwood system has been widely used to create regeneration layers (Malcolm et al. 2001; Wetzel 2001). In this silvicultural system, shade-tolerant species and opportunistic species are used for enrichment plantings because they survive and develop well under a forest canopy (Wang 2018). These shade-tolerant species and opportunistic species include M. rigida, C. glauca, A. japonicum, C. fargesii, M. pauhoi, V. Montana, and L. glaber. Considering the species associations, the following species pairs are recommended: A. japonicum (U) - M. rigida (U) and A. japonicum (U) - L. glaber (U). Of course, after shelterwood cutting, the positively associated pairs of shade-intolerant species can be replanted in relatively large open areas, e.g., S. discolor (U) - E. roxburghiana (U).
Additionally, O'Hara (2014) demonstrated that creating forest gaps is a suitable approach for rapidly establishing a regeneration layer. Due to good light conditions in forest gaps, shade-intolerant species and opportunistic species can survive and grow (Wang 2018). We therefore suggest the following shade-intolerant species and opportunistic species: S. discolor, E. roxburghiana, L. mollis, M. pauhoi, V. montana, L. glaber, and C. fargesii. The final species pairs that were determined in this study included S. discolor (U) - E. roxburghiana (U), V. montana (S) - L. mollis (S), and M. pauhoi (U) - S. discolor (U). Furthermore, we should stress that negatively associated tree species are not encouraged to be planted together, e.g., M. rigida (S) - L. mollis (S) and C. fargesii (S) - C. glauca (S), though positive complementary effects might exist. Additionally, in order to accelerate natural succession, high-value shade-tolerant species and opportunistic species such as C. fargesii, L. glaber, and V. montana can be planted in the understory without having to create forest gaps.
The main canopy of the most degraded natural forests is degenerated and hence the facilitation of tree regeneration is of great importance for successful restoration. However, due to forest fragmentation, the seed dispersal mechanism is not functioning properly, resulting in a lack of natural regeneration of high-value and late-successional tree species (Wang 2018; Aide et al. 2000). Therefore, it is necessary to plant positively associated, high-value, and late-successional tree species in degraded natural forests to promote regeneration. The following positively associated shade-intolerant species or opportunistic species are recommended to fill natural forest gaps: S. discolor (U) - E. roxburghiana (U), V. montana (S) - L. mollis (S), and M. pauhoi (U) - S. discolor (U). Although it is not always necessary to create forest gaps in natural forests, it might be required in dense parts of the forest.
In the understory, the saplings of shade-tolerant species or opportunistic species that are positively associated with the nearby adult tree species are recommended for under-planting. These pairs include montana (S) - L. mollis (U), montana (S) -montana (U), M. pauhoi (S) - E. roxburghiana (U), M. rigida (S) - L. glaber (U), M. rigida (S) - A. japonicum (U), M. rigida (S) - M. rigida (U), M. rigida (S) - S. discolor (U), M. rigida (S) - M. pauhoi (U), C. glauca (S) - E. roxburghiana (U), C. glauca (S) - M. pauhoi (U), C. glauca (S) - S. discolor (U), V. montana (S) - A. japonicum (U), and C. fargesii (S) - C. fargesii (U). It is suggested that the under-planted saplings should have a positive association with other saplings.
Additionally, it is also necessary to consider the relationship between the under-planted tree species and the nearby adult tree species and avoid planting negatively associated species pairs. For example, it is suggested to avoid planting the saplings of M. rigida under the adult trees of L. mollis, the saplings of A. japonicum under the adult trees of S. discolor, and the saplings of C. fargesii under the adult trees of M. pauhoi. Moreover, sapling pairs with a significant negative association should also be avoided, e.g., M. rigida (S) - L. mollis (S) and C. fargesii (S) - C. glauca (S).
4.4 Potential shortcomings of this study
The species interactions occur within a spatial scale beyond which they would no longer exist (Velázquez et al. 2015; Jonsson 1998; Guo et al. 1997). Therefore, the suitable number of sample plots and plot size for examining species association should be determined according to the species or vegetation characteristics (Velázquez et al. 2015; Schulz et al. 2009; Zhao et al. 2003; Margóczi 1995). According to the current literature, our plot size is close to the ideal size of subtropical evergreen broad-leaved forest in southern China (Zhou 2000; Guo et al. 1997; Du et al. 1995, Wang 1985).Furthermore, Xu (2016) argued that if the ratio of species pairs of positive association to the species pairs of negative associations was equal or close to 1, the field survey design, which included the number of sample plots and plot size, was promising and could accurately capture the species association. In the present study, the ratio of the species pairs of positive association to species pairs of the negative association was close to 1.
Biased estimation may occur if a chi-square test is performed using small sample plot (Zar 1974). Therefore, we performed Yate's correction to remove bias; this method has been extensively used by many ecologists when investigating species association with small sample plot (Matchima et al. 2018; Zhao et al. 2018; Gu et al.,2017; Zhao et al.,2012; Ludwig et al. 1988). Additionally, there is no specific requirement for the sample size for the AC, DI, and SRCC, though a minimum of two variables with at least 8 to 10 observations for each variable is recommended for the calculating SRCC. Therefore, we concluded that the species association derived from the 2500 m2 sample plot was reliable.
5. Conclusions
In this study, we investigated the intra- and interspecies spatial associations between saplings and adult trees in an old-growth forest in western Hunan Province. We considered the light requirements of the tree species and the optimal species configuration to provide suggestions for silvicultural practices for the establishment of mixed-species plantations through afforestation, transforming monocultures, and enrichment plantings in degraded secondary natural forests. The results of our study provide a theoretical basis for sustainable forest management in western Hunan Province.
Acknowledgements
We thank our colleagues in Huitong National Research Station of Forest Ecosystem for their support during our field work. This research was funded by the National Key R&D Program of China, grant number 2017YFC0505604.
References
Adams T, Ackland G, Marion G, and Edwards C. 2011. Understanding plantation transformation using a size-structured spatial population model. Forest Ecology and Management 261:799-809.
Ådjers G, Hadengganan S, Kuusipalo J, Nuryanto K, and Vesa L. 1995. Enrichment planting of dipterocarps in logged-over secondary forests: effect of width, direction and maintenance method of planting line on selected Shorea species. Forest Ecology and Management 73:259-270.
Aide T, Zimmerman J, Pascarella J, Rivera L, and Marcano‐Vega H. 2000. Forest regeneration in a chronosequence of tropical abandoned pastures: implications for restoration ecology. Restoration ecology 8:328-338.
Amoroso M, and Turnblom E. 2006. Comparing productivity of pure and mixed Douglas-fir and western hemlock plantations in the Pacific Northwest. Canadian Journal of Forest Research 36:1484-1496.
Bruijnzeel L. 2004. Hydrological functions of tropical forests: not seeing the soil for the trees? Agriculture, ecosystems & environment 104:185-228.
Cai N, Li G, and Lu Y. 2006. Discussion on the Approaching-nature Forestry Mangement of Pinus yunnanensis Pure Forests. Journal of Northwest Forestry University 21:85.
Camilli G, and Hopkins K. 1978. Applicability of chi-square to 2× 2 contingency tables with small expected cell frequencies. Psychological Bulletin 85:163-167.
Chai Z, Sun C, Wang D, and Liu W. 2016. Interspecific associations of dominant tree populations in a virgin old-growth oak forest in the Qinling Mountains, China. Botanical studies 57:23.
Cole, and Lamont C. 1949. The measurement of interspecific associaton. Ecology 30:411-424.
De Groot R, Wilson M, and Boumans R. 2002. A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecological economics 41:393-408.
Deng L, Bai X, Li L, Niu S, Han M, Qin S, and Zhou Y. 2015. Interspecific association and correlation among dominant woody plants of secondary forest in montane region of eastern Liaoning Province, China. Chinese Journal of Applied Ecology 34:1473-1479.
Deng Q, Liu S, Liu J, Meng Z, and Zhang D. 2007. Contributions of litter-fall to soil respiration and its affacting factors in southern subtropical forests of China. Advances in Earth Science 22:976-986.
Dezfuli B, Giari L, De Biaggi S, and Poulin R. 2001. Associations and interactions among intestinal helminths of the brown trout, Salmo trutta, in northern Italy. Journal of Helminthology 75:331-336.
Du D, Liu Y, and Li R. 1995. Studies on the interspecific association of dominant species in a subtropical Catanopsis fargesii forest of Jinyun Mountain, China. Acta Phytoecol Sin, 19:149-157.
Du L, Yang H, Guo X, Dong D, and Guan W. 2013a. Interspecific associations of Pistacia chinensis community and association index-rank pattern model. Journal of Beijing Forestry University 35:37-45.
Du M, Ding G, and Cai Q. 2018. The Transcriptomic Responses of Pinus massoniana to Drought Stress. Forests 9:326.
Du Z, Kang X, Meng J, Kong L, Guo W, and Yue G. 2013b. Spatial Distribution Pattern and Association of Dominant Tree Species in Poplar-Birch Secondary Forest Stand in Changbai Mountains. Journal of Northeast Forestry University 41:36-42.
Erman D, and Helm W. 1971. Comparison of some species importance values and ordination techniques used to analyse benthic invertebrate communities. Oikos 22:240-247.
Felton A, Lindbladh M, Brunet J, and Fritz Ö. 2010. Replacing coniferous monocultures with mixed-species production stands: an assessment of the potential benefits for forest biodiversity in northern Europe. Forest Ecology and Management 260:939-947.
Felton A, Nilsson U, Sonesson J, Felton A, Roberge J, Ranius T, Ahlström M, Bergh J, Björkman C, and Boberg J. 2016. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 45:124-139.
Gautheir T. 2001. Detecting trends using spearman's rank correlation coefficient. Environmental forensics 2:359-362.
Greig-Smith P. 1983. Quantitative Plant Ecology (3rd ed. ) . Oxford: Blackwell Scientific Publications.
Gu L, Gong Z, and Li W. 2017. Niches and Interspecific Associations of Dominant Populations in Three Changed Stages of Natural Secondary Forests on Loess Plateau, PR China. Scientific reports 7:6604.
Guo Z, Zhuo Z, Chen J, and Wu M. 1997. Interspecific Association of Trees in Mixed Evergreen and Deciduous Broadleaved Forest in Lushan Mountain. Acta Phytoecologica Sinica 21:424-342.
Hansen A, McComb W, Vega R, Raphael M, and Hunter M. 1995. Bird habitat relationships in natural and managed forests in the west Cascades of Oregon. Ecological applications 5:555-569.
Hao Z, Zhang J, Song B, Ye J, and Li B. 2007. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. Forest Ecology and Management 252:1-11.
He Y, Qin L, Li Z, Liang X, Shao M, and Tan L. 2013. Carbon storage capacity of monoculture and mixed-species plantations in subtropical China. Forest Ecology and Management 295:193-198.
Hou J, Mi X, Liu C, and Ma K. 2004. Spatial patterns and associations in a Quercus‐Betula forest in northern China. Journal of Vegetation Science 15:407-414.
Hou J, Yin R, and Wu W. 2019. Intensifying Forest Management in China: What does it mean, why, and how? Forest Policy and Economics 98:82-89.
Hu W, Huang S, Li J, Zhou G, and Wang X. 2013. Interspecific associations among dominant tree populations in an elfin forest community in Shikengkong of Nanling Mountains, China. Chinese Journal of Ecology 32:2665-2671.
Huang Y, Chen Y, Castro-Izaguirre N, Baruffol M, Brezzi M, Lang A, Li Y, Härdtle W, von Oheimb G, and Yang X. 2018. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 362:80-83.
Hurlbert S. 1969. A coefficient of interspecific assciation. Ecology 50:1-9.
Isango J, Varmola M, Valkonen S, and Tapaninen S. 2007. Stand structure and tree species composition of Tanzania miombo woodlands: a case study from miombo woodlands of community based forest management in Iringa district. Proceedings of the 1st MITIMIOMBO Project Workshop 50:43-56.
Jian M, Liu Q, Zhu D, and You H. 2009. Inter-specific correlations among dominant populations of tree layer species in evergreen broad-leaved forest in Jiulianshan Mountain of subtropical China. Journal of Plant Ecology 33:672-680.
Jiang Y, Chen C, Liu Y, and Xu Z. 2010. Soil soluble organic carbon and nitrogen pools under mono-and mixed species forest ecosystems in subtropical China. Journal of Soils and Sediments 10:1071-1081.
Jonsson B., and Moen J. 1998. Patterns in species associations in plant communities: the importance of scale. Journal of Vegetation science 9:327-332.
Kahmen A, Renker C, Unsicker S, and Buchmann N. 2006. Niche complementarity for nitrogen: an explanation for the biodiversity and ecosystem functioning relationship? Ecology 87:1244-1255.
Kimmins J. 2003. Old-growth forest: An ancient and stable sylvan equilibrium, or a relatively transitory ecosystem condition that offers people a visual and emotional feast? Answer it depends. The Forestry Chronicle 79:429-440.
Klopčič M, Simončič T, and Bončina A. 2015. Comparison of regeneration and recruitment of shade-tolerant and light-demanding tree species in mixed uneven-aged forests: experiences from the Dinaric region. Forestry: An International Journal of Forest Research 88:552-563.
Legendre L,and Legendre P. 1978. Associations. Phytoplankton manual 6:261–272.
Li J, Xie F, Chen C, Zhang S, Xiao R, and Zhang D. 2001. Interspecific association of dominant species in Betula luminifera natural forest communities of Shaowu, Fujian Province. Chinese Journal of Applied Ecology 12:168-170.
Li W. 2004. Degradation and restoration of forest ecosystems in China. Forest Ecology and Management 201:33-41.
Li Y, Yang L, Ji Y, Sun H, Li H, and Wang W. 2008. Lead uptake by plant in soil-plant system at lead-zinc deposit area of Western Hunan Province. Huan jing ke xue= Huanjing kexue 29:196-201.
Lin D, Xian D, Xing S, Gui G, and Zhang W. 2008. Interspecific association analysis in dominant species of Juglans mandshurica community in Wuling Mountain Nature Reserve of Beijing. Journal of Beijing Forestry University 30:154-158.
Lin Y, Wu C, Hong W, Ji G, Hu X, and Wu J.2005. Study on the scale effect of interspecific association of species in tree layer of the rare plant Tsuga longibracteata community. Guihaia 25:526-532.
Liu H, Du W, Wang Y, Chen Y, Wu Y, and Yuan l. 2017. Effects of Eupatorium adenophorum on interspecific association and the stability of companion species in Liangshan Prefecture of Sichuan Province. Acta Ecologica Sinica 37:5031-5038.
Liu Q, Kang M, and Liu Q. 2006. Quantitative classification and environmental interpretation of forest tree species in Hungou, Zhongtiao Mountain. Acta Phytoecological Sinica 30:383-391.
Ludwig J, Quartet L, and Reynolds J. 1988. Statistical ecology: a primer in methods and computing. John Wiley & Sons.
Luo Y, Sun D, Lin J, and Guo W. 2013. Effect of Close-to-Nature Mangement on the natural regeneration and species diversity in a masson pine plantation. Acta Ecologica Sinica 33:6154-6162.
Luo Z, Yu M, Chen D, Wu Y, and Ding B. 2012. Spatial associations of tree species in a subtropical evergreen broad-leaved forest. Journal of Plant Ecology 5:346-355.
Malcolm D, Mason W, and Clarke G. 2001. The transformation of conifer forests in Britain-regeneration, gap size and silvicultural systems. Forest Ecology and Management 151:7-23.
Manson D, Hanan J, Hunt M, Bristow M., Erskine P, Lamb D, and Schmidt S. 2006. Modelling predicts positive and negative interactions between three Australian tropical tree species in monoculture and binary mixture. Forest ecology and management, 233:315-323.
Mangueira J, Holl K, and Rodrigues R. 2019. Enrichment planting to restore degraded tropical forest fragments in Brazil. Ecosystems and People 15:3-10.
Margóczi K. 1995. Interspecific associations in different successional stages of the vegetation in a Hungarian sandy area. Tiscia 29:19-26.
Matchima K, Vongprasert J, and Chutiman N. 2018. The Development of a Correction Method for Ensuring a Continuity Value of The Chi-square Test with a Small Expected Cell Frequency. Naresuan University Journal: Science and Technology 26:98-105.
Meng J, Lu Y, and Zeng J. 2014. Transformation of a degraded Pinus massoniana plantation into a mixed-species irregular forest: impacts on stand structure and growth in southern China. Forests 5:3199-3221.
Mundry R, and Fischer J. 1998. Use of statistical programs for nonparametric tests of small samples often leads to incorrect Pvalues: examples from Animal Behaviour. Animal behaviour 56:256-259.
Ning J, Lu Y, Zhao H, Liu X, Ren Y, and Chen J. 2009. Assessment on close-to-nature transformation of Pinus tabulaeformis plantations in Xishan region, Beijing. Journal of Northeast Forestry University 37:42-44.
O'Hara K. 1998. Silviculture for structural diversity: a new look at multiaged systems. Journal of forestry 96:4-10.
O'Hara K. 2014. Multiaged silviculture: managing for complex forest stand structures: Oxford University Press, USA.
O’Hara K. 2001. The silviculture of transformation—a commentary. Forest Ecology and Management 151:81-86.
Peng S, Zhou H, Guo S, and Huang Z. 1999. Studies on the changes in interspecific association of zonal vegetation in dinhushan. Acta botanica sinica 41:1239-1244.
Pimentel D, Stachow U, Takacs DA, Brubaker HW, Dumas AR, Meaney JJ, Onsi DE, and Corzilius DB. 1992. Conserving biological diversity in agricultural/forestry systems. BioScience 42:354-362.
Ramos J, and del Amo S. 1992. Enrichment planting in a tropical secondary forest in Veracruz, Mexico. Forest Ecology and Management 54:289-304.
Rhoades H, and Overall J. 1982. A sample size correction for Pearson chi-square in 2× 2 contingency tables. Psychological Bulletin 91:418.
Richards A, Forrester D, Bauhus J, and Scherer-Lorenzen M. 2010. The influence of mixed tree plantations on the nutrition of individual species: a review. Tree physiology 30:1192-1208.
Schluter D. 1984. A variance test for detecting species associations, with some example applications. Ecology 65:998-1005.
Schulz, B, Bechtold W , and Zarnoch S. 2009. Sampling and estimation procedures for the vegetation diversity and structure indicator. US Department of Agriculture, Forest Service, Pacific Northwest Research Station.
Spies T, and Franklin J. 1996. The diversity and maintenance of old-growth forests. Biodiversity in managed landscapes Oxford University Press, New York, New York, USA 296-314.
Su S, Liu J, He Z, Zheng S, Hong W, and Xu D. 2015. Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain National Nature Reserve. Journal of Mountain Science 12:637-646.
Sun D, Wen Y, Luo Y, and Li X. 2015. Effec of Close-to-Nature Management on Species Diversity in a Cunninghamia lanceolata Plantation. Forest Research 28:22.
Sushma H, and Singh M. 2006. Resource partitioning and interspecific interactions among sympatric rain forest arboreal mammals of the Western Ghats, India. Behavioral Ecology 17:479-490.
Thomas J Ruggiero L, Mannan R, Schoen J, and Lancia R. 1988. Management and conservation of old-growth forests in the United States. Wildlife Society Bulletin (1973-2006) 16:252-262.
Tigabu M, Savadogo P, Odén PC, and Xayvongsa L. 2010. Enrichment planting in a logged-over tropical mixed deciduous forest of Laos. Journal of Forestry Research 21:273-280.
Van Couwenberghe R, Gégout J-C, Lacombe E, and Collet C. 2013. Light and competition gradients fail to explain the coexistence of shade-tolerant Fagus sylvatica and shade-intermediate Quercus petraea seedlings. Annals of botany 112:1421-1430.
Velázquez C, Eduardo E, Paine T, May, F, Wiegand T , and De Bello F. 2015. Linking trait similarity to interspecific spatial associations in a moist tropical forest. Journal of Vegetation Science, 26:1068-1079.
Wang B, and Peng S. 1985. The measuring technique of interspecific association of the lower-subtropical evergreen -broadleaf forest II. The sample echnique tropical and subtropical forest ecosystem 3:167-173.
Wang H, Huang Y, Feng Z, and Wang S. 2007. C and N stocks under three plantation forest ecosystems of Chinese fir, Michelia macclurei and their mixture. Frontiers of Forestry in China 2:251-259.
Wang H, Liu S, Wang J, Shi Z, Lu L, Zeng J, Ming A, Tang J, and Yu H. 2013. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. Forest Ecology and Management 300:4-13.
Wang J, and Meng J. 2018. Identifying indigenous tree species for land reforestation, forest restoration, and plantation transformation on Hainan Island, China. Journal of Mountain Science 15:2433-2444.
Wang Q, Bao D, Guo Y, Lu J, Lu Z, Xu Y, Zhang K, Liu H, Meng H, and Jiang M. 2014. Species associations in a species-rich subtropical forest were not well-explained by stochastic geometry of biodiversity. PloS one 9:97300.
Wang Q, Wang S, and Huang Y. 2008. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. Forest Ecology and Management 255:1210-1218.
Wang X, Feng Z, and Ouyang Z. 2001. The impact of human disturbance on vegetative carbon storage in forest ecosystems in China. Forest Ecology and Management 148:117-123.
Wetzel S, and Burgess D. 2001. Understorey environment and vegetation response after partial cutting and site preparation in Pinus strobus L. stands. Forest Ecology and Management 151:43-59.
White DL, and Lloyd FT. 1994. Defining old growth: Implications for management. USDA Forest Service.
Wiegand T, Gunatilleke S, and Gunatilleke N. 2007. Species associations in a heterogeneous Sri Lankan dipterocarp forest. The American Naturalist 170:77-95.
Wilson E, Short I, Ní Dhubháin Á, and Purser P. 2018. Transforming Sitka spruce plantations to continuous cover forestry. Forestry & Energy Review 8:38-40.
Xu M, Liu M, Zhai D, and Liu T. 2016. A review of methods used to analyzje various aspects of plant interspecific associations. Acta Ecologica Sinica 36.
Xue O, and Wei T. 2016. Interspecific Association among Understory species of the Low-efficiency Plantation in the Jiufeng national Forest Park. Bulletin of Botanical Research 36:34-42.
Ye N, Shen N, Shang T, Gao H, Guan J, and Yi L. 2017. Vegetation Structure and Internal Relationship Between Distribution Patterns of Vegetation and Environment in Ecological Service Forest of Rui'an City in Zhejiang Province. Bulletin of Botany 52:496-510.
Xiong Y, Zeng G, Chen G, Tang L, Wang K, and Huang D. 2007. Combining AHP with GIS in synthetic evaluation of eco-environment quality – a case study of Hunan Province, China. Ecological modelling 209:97-109.
Yuan D, Lin L, Yan C, Wu Y, Liu Q, and Zhou Z. 2012. Afforestation Effect of Three Species of Seedlings Growing with Light Media in Net Container. Journal of Northeast Forestry University 4:19-23.
Yuan Z, Wei B, Chen Y, Jia H, Wei Q, and Ye Y. 2018. How do similarities in spatial distributions and interspecific associations affect the coexistence of Quercus species in the Baotianman National Nature Reserve, Henan, China. Ecology and evolution 8:2580-2593.
Yang Y. 2001. Impacts and effectiveness of logging bans in natural forests: People's Republic of China. Forests out of bounds: impacts and effectiveness of logging bans in natural forests in Asia–Pacific Food and Agricultural Office of the United Nations (FAO), Bangkok:81-102.
Zar G. 1974. Biostatistical statistical analysis. Englewood Cliffs, NJ, Prentice-Hall Inc.
Zhang J, Cheng K, Zang R, and Lang N. 2011. Construction of naturalness evaluation index system for different communities of Pinus yuannanensis forest. Journal of Agriculture University of Hebei 34:71-76.
Zhang R, Guo L, Yi X, Cao W, Wang Y, Wu P, and Ji L. 2015. Analysis of interspecific associations among major tree species in three forest communities on the north slope of Changbai Mountain. Acta Ecologica Sinica 35:106-115.
Zhang X Zhang X, Guo C, and Ding J. 2012. Quantitative dynamics of Pteroceltis tatarinowii populations in Huangcangyu Nature Reserve of Anhui Province, East China. Chinese Journal of Ecology 31:282-291.
Zhao H, Kang X, Guo Z, Yang H, and Xu M. 2012. Species Interactions in Spruce–Fir Mixed Stands and Implications for Enrichment Planting in the Changbai Mountains, China. Mountain Research and Development 32:187-196.
Zhao Y, Jin H, Huang L, Chen Q, Liu L, Dai Y, and Wang C. 2018. Spatial Distribution Pattern and Interspecific Association Analysis of Thuja koraiensis Population. E3S Web of Conferences 53.
Zhao, Z., Zu, Y., Yang, F., and Cong, P. 2003. Study on the sampling technique of interspecific association of ligneous plant in Quercus liaotungensis forest in Dongling Mountain. Acta Phytoecological Sinica 27: 396-403.
Zhou L, Yu H, and Chen K. 2002. Relationship between microcystin in drinking water and colorectal cancer. Biomed Environ Sci 15:166-171.
Zhou X, Wang B, Li M, and Zan Q. 2000. An analysis of interspecific associations in secondary succession forest communities in Heishiding Nature Reserve, Guangdong Province. Acta Phytoecologica Sinica, 24:332-339.



