Schlüsselbegriffe: Ellenberg´scher Indikator L-Wert; Lochhieb; Mittelwald; Carpinus betulus
Abstract
The aim of this study was to document species composition and richness as well as light affinity of the herbaceous understorey in an oak-hornbeam coppice-with-standards forest with respect to phytosociology. Our research questions were (i) how does the herbaceous understorey react to interventions of different size, (ii) how do their light indicators values (Ellenberg et al. 2001) reflect silvicultural methods, and (iii) do local temperatures mirror the respective light environment? In an oak-hornbeam coppice-with-standards forest in North-eastern Lower Austria with the most recent coppice harvesting in 1958, the effects of two silvicultural methods on the herbaceous layer were investigated. The treatments were (i) a coppice with standards treatment for 20 years, and ii) small gap clearings for 7 years. Species richness and composition as well as indicator values were compared to the surrounding forest, since the small gap-clearings were also coppiced in 2009. Phytosociological relevés and the calculation of light indicator values indicated that small gap clearings appear to be more effective in re-establishing an understorey community close to the typical forest community than coppicing. One reason is that small gaps provide light conditions for the understorey vegetation similar to the forest. More light-affine species cannot easily penetrate into these gaps from outside the forest. Very dense Carpinion stands shifting towards other Fagetalia communities should be coppiced more often, but on a small scale to promote herbaceous Carpinion species in the understorey. Thus forest management can help preserve these site-specific typical vegetation communities.
Introduction
Many parts of Europe provide the climatic and pedological environment for oak-hornbeam forests. These forests cover low elevations that are easily accessible. For centuries they have produced quality timber as well as heating material and food. Near settlements, they also often served as pastures (Starlinger 1997). Some of these forests, in turn, may have developed after abandonment of arable fields (Laska 1997). Until the late 19th century, the fast-growing hornbeam (Carpinus betulus) was more often and on smaller scales harvested than oaks; thus stand structure and conditions for the Carpinion understorey were kept more favorable. After this period the former coppice-with-standards system has been changed for economic reasons (Jahn & Raben 1982), and many of these forests were converted into high forests. Often the natural canopy was replaced by planted, fast-growing or economically promising tree species, such as Scots pine (Pinus sylvestris) in a Pannonian steppe forest (Chudomelová et al. 2017), or the frequency of coppicing was partially drastically reduced (Hédl et al. 2010; Becker et al. 2017). There is plenty evidence, that changes in light availability are an important driving factor in successions (Faliński 2003; Axmanová et al. 2012; Jamrichová et al. 2013; Bernhardt-Römermann et al. 2015). Light promotes Carpinion species regeneration and allows endangered light-loving species to survive (Roleček et al. 2017). This has been well documented for dense forests in Great Britain, where thinning enhanced species richness and the development towards high forests reduced it (Kirby et al. 2017a; Kirby et al. 2017b). Nutrient accumulation (nitrogen, in particular) in the soils of recently developed high forests is discussed controversially in literature (Becker et al. 2017). Higher average temperatures, less and irregular precipitation, air pollution, and biological pests such as oak mistletoe, or high numbers of deer and wild boar led to a discussion on how to maintain the productivity of forests as carbon sinks and renewable resources (Bruckman et al. 2016; Gerhardt et al. 2013; Hagen 2005; Laurent et al. 2017). Gilliam (2007) showed that the herbaceous layer is of crucial importance for secondary successions in temperate forests. The immigration of light-affine non-native species, fostered by anthropogenic seed transport, has also been discussed in this context (Chytrý et al. 2009; Kleinbauer et al. 2010; Essl et al. 2011; Pyšek & Chytrý 2014). The import of non-forest seeds by wind and humans into large coppice or clearcuttings areas reduces the likelihood of regeneration from the local soil seed bank, as forest species usually have low competitive power and lower light demand. Frost & Rydin (1997) stated that oak seedlings may be outcompeted by dense herbaceous ground vegetation. Especially light-affine non-native species appear to hinder the re-establishment of site-specific forest vegetation. In the past few decades, parasites, diseases and extreme weather conditions resulted in increasing economic pressure on forest managers and to mitigate the associated effects various silvicultural methods have been tested with the goal of restoring diversity and promoting rare and endangered forest understorey plants.
This applied also to our study site in a Pannonian oak-hornbeam forest and there had been no forest management for almost 40 years until 2009 (Hochbichler 2008). We aim to answer the following questions:
1.) How do herbaceous species richness and composition react to interventions of different quality and intensity?,
2.) do light indicator values (Ellenberg et al. 2001) reflect management changes, and
3.) do local temperature mirror the respective light environment?
Methods
Study area
This research has been conducted in a fenced oak-hornbeam coppice-with-standards forest not coppiced since 1958 at 48°24´55´´N, 16°35´50´´E, in NE Lower Austria. Hochbichler (2008) reports, that the study site has been formerly used as arable land, before the forest was established some hundred years ago. The study area is situated in the Pannonian region on a slightly inclined plateau, oriented SW to NE. The forest soils in this region hosting Quercus petraea and Carpinus betulus are predominantly nutrient rich cernosems (Starlinger 1997, www.bfw.ac.at/ebod, last access 2019-04-30). Ulrich (1987) described the soils as loamy sands with a moderate and variable water holding capacity. From the phytosociological point of view, the forest resembles the locally dominating Galio sylvatici-Carpinetum (Willner & Grabherr 2007), but the moderately light-affine Carpinion and Galio-Carpinetum species (Galium sylvaticum or Lathyrus niger) generally reach lower coverage than the mor frequent, shade tolerant Fagetalia species such as Polygonatum multiflorum or Galium odoratum. As described at Hochbichler (2008), the canopy of the forest was composed by Quercus petraea (about one individual per 600 m²) and a few individuals of Quercus cerris, Sorbus domestica and Sorbus torminalis. Carpinus betulus and Acer campestre form a dense mid-storey. Detailed silvicultural information including stand structure and growth during the experiment are given in Hochbichler (2008).

Figure 1: Aerial photo (oriented) of the study site, year 2000 (Bundesamt für Eich- und Vermessungswesen, Vienna, Austria). / Abbildung 1: Luftbild (gerichtet) der Versuchsflächen aus dem Jahr 2000 (Bundesamt für Eich- und Vermessungswesen, Wien, Österreich).
The Pannonian climate registered 1971-2000 at Hohenau/March (about 30 km NE) was characterized by a mean maximum temperature of 26°C (absolute maximum of 37 °C) in August. In this period the yearly average temperature was 9.3 °C and the annual precipitation sum was 498 mm. Average temperatures for 1961-1990 at Hohe Warte, Vienna (40 km S) was 10.3 °C, which was by 1.1 °C higher than the average temperature in the period 1871-1900. A similar rise of temperature for the study area appear likely (www.zamg.ac.at, last access 2019-04-30).
Study design
The almost square total area (about 3.5 ha) had been divided into 25 m² plots by a 5 by 5 m grid (Hochbichler, 2008). Table 1 gives a description of the permanent sampling plots. The locations of the differently treated study sites labelled as “forest”, “large coppice with standards”, “small gap clearings”, and “forest plots adjacent to small gap clearings” are shown in Figure 1. “Forest” represents an unthinned stand. The “large coppice with standards” had a size of 1.5 ha and before coppicing this area was part of the homogeneous forest area. For every “small gap clearing” the wood biomass (trees and shrubs) within a 15 by 15 m plot were completely removed. The 2 ha of “forest” in the western part were coppiced in 2009 due to usual management, including all plots of the “small gap clearings” and the “forest plots adjacent to small gap clearings”. In turn, no further inventory was possible except in the “large coppice with standards” (Table 1).
Hochbichler (2008) reported increasing canopy cover in the old coppice-with-standards forest from 80 % to 95 % (1996 to 2002) as a consequence of the vigorous regrowth in the lower layers. Diameters of the oaks amounted to > 18 cm up to > 50 cm in the old coppice-with-standards forest. In the “large coppice with standards”, the tree layer covered 20 % - 30 %.
Shaded mechanical thermo-hydrographs recorded air temperature 2 m above ground in the vegetation periods from April to September in 1999 and 2000 in the three types of treatments – “forest”, “small gap clearings” and “large coppice with standards”. Missing records in figure 2 were due to mistakes and failures of the devices. Averaged values from Vienna were calculated for the same periods.
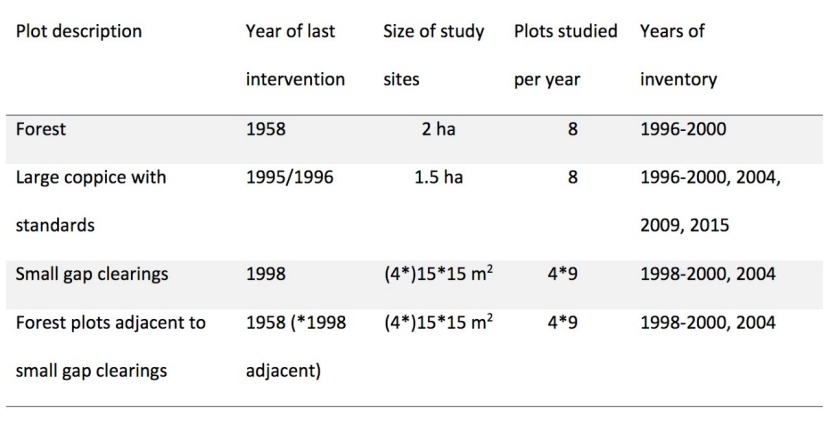
Table 1: Silvicultural treatments, years of last intervention, size of study sites, number of relevé plots per year and site, and years of inventory. / Tabelle 1: Waldbauliche Maßnahmen, Zeitpunkt der letzten Eingriffe, Flächengrößen, Anzahl der Aufnahmeflächen pro Jahr und Maßnahmenfläche, Jahre der Aufnahmen.
Vegetation data including statistical analyzes
All plots were investigated repeatedly using the Londo (1976) method. For all relevés, single species cover was noted in the ground layer as well in the tree and shrub layers. The relevés inform about the estimated horizontal projection of ground cover by every plant species. Cumulated cover was calculated for every layer as well as for the whole community. Herbaceous forest species were grouped according to their affiliation to phytosociological units (Willner & Grabherr 2007); non-forest plants (e.g. clearcuts, ruderal, segetal places or turfs/meadows) according to Oberdorfer (2001). Nomenclature of all species follows Fischer et al. (2008).
Species coverage data were used to calculate weighted indicator values adapted to Austrian conditions (Ellenberg et al. 2001; Karrer & Kilian 1990). Tree, shrub, and ground layers were treated separately. Significant differences were tested by means of ANOVA and Scheffé as a post hoc test in the program SPSS 21 (® IBM) on a significance level of α = 5 %. Weighted indicator values were tested using non-parametric Kruskal-Wallis-test and Bonferroni-Dunn as post-hoc test with the same significance level.
Results
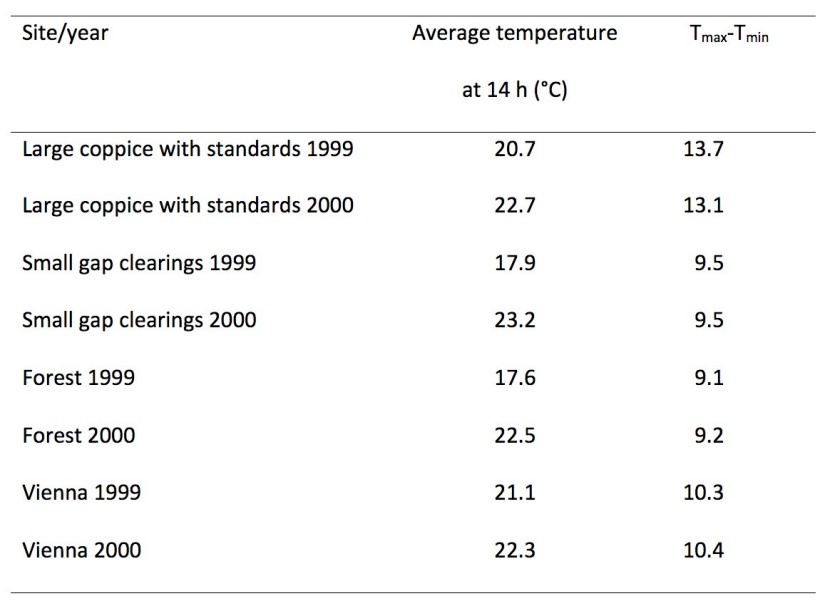
Table 2: Average values of registered temperatures with gaps in the recordings –see figure 2 (1st April to 30th September). / Tabelle 2: Mittlere aufgezeichnete Temperaturen mit Lücken im Datensatz – siehe Abbildung 2 (1. April bis 30. September).
For the six month period from 1st April to 30th September the precipitation sum in Vienna amounted to 438 mm in 1999, but only 250 mm in 2000 (www.zamg.ac.at, last access 2019-04-30). The forest and the small gap clearings provided a more moderate temperature regime than the large coppice with standards (Table 2). The largest differences between extreme temperature values were measured in the large coppice. In the small gap clearings and the forest the differences were significantly lower (Figure 2).
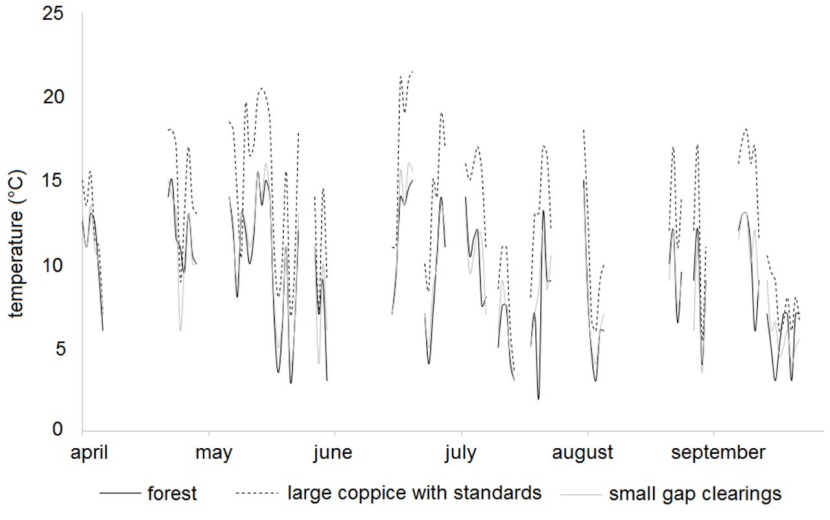
Figure 2: Differences between daily minimum and maximum temperatures for summer 2000, recorded for forest, large coppice with standards, and southern small gap clearings. / Abbildung 2: Unterschiede zwischen täglichem Minimal- und Maximaltemperaturen im Sommer 2000, aufgenommen in Wald, großer Schlagfläche mit Überhältern und dem südlichen Lochhieb.
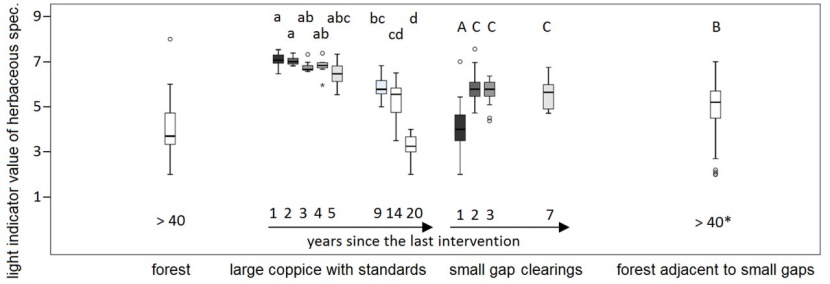
Figure 3: Average weighted L- values for herbaceous species in the plot groups forest, large coppice with standards, small gap clearings, and forested plots adjacent to small gaps. *later intervention in adjacent gaps! abcd and ABC refer to significantly different groups (Bonferroni-Dunn test, α=0.05), see text. / Abbildung 3: Mittlere gewichtete L-Werte für die Krautvegetation in den Aufnahmegruppen Wald, großer Schlag mit Überhältern, kleine Lochhiebe, und an die Lochhiebe angrenzender Wald. *späterer Eingriff in den benachbarten Flächen! abcd und ABC beziehen sich auf signifikant unterschiedliche Gruppen (Bonferroni-Dunn test, α=0.05), siehe Text.
Light indicator values
The high irradiation on the large coppice with standards is also reflected by the temperature records (Table 2 and Figure 2) and the light indicator values of herbaceous species (Figure 3). Carpinion herbs/ forbs usually have Ellenberg indicator L-values (Ellenberg et al. 2001; Karrer & Kilian 1990) of four to six (on a scale from one – deep shade – to nine – full sun light). In the large coppice, L-values easily reach levels above seven in the first decade. 20 years after coppicing, the L-value of the herb layer in the newly grown forest in the coppice with standards treatment is significantly lower than the old forest. The L-values of the small forested plots adjacent to the gaps are significantly lower than those in the gaps themselves, but significantly higher than in the dense coppice with standards after 20 years and the old, unthinned forest.
The small gaps start at a level comparable to the forest. Obviously, the forest plant community can outlive the first year after the intervention. The years three and seven after the last intervention show higher portions of Carpinion/ Galio-Carpinetum species with L-values between five and six both in gaps and adjacent forest plots, and a portion of non-forest species with high L-values. Whereas the L-value of herbaceous species does not differ significantly between years in the adjacent forest plots (results not shown), it rises significantly in the small gap clearings from year one to year two, but stays at a lower level than in the large coppice.
All other Ellenberg indicator values (temperature, continentality, moisture, nutrients and soil reaction) were evaluated in the same way as light indicator values. The latter proved to be the best to mirror the vegetation dynamics, however. Therefore only these results are presented and discussed in detail. Especially temperature and nutrients followed an expectable pattern but significances were more overlayed by random effects and higher numbers of species with no specific value for these variables compared to L-values.
Species numbers and coverage
Fig. 4 shows low numbers of non-forest species (Table A1) in forest and forested plots adjacent to small gaps, but extremely high ones in the first three years after the coppice with standards in the respective plots. Nine to fourteen years after the coppice, these species numbers shrink almost to the forest level. The maximum number of non-forest species penetrated into the small gaps with a time lag of one year. The number in the gaps amounts to only about 2/3 of the number reached in the large coppice with standards.

Figure 4: Number of non-forest species (i.e. clearcut, ruderal, segetal and turf/ meadow species) in the plot groups forest, large coppice with standards, small gap clearings, and forested plots adjacent to small gaps. * later intervention in adjacent gaps! abcd and ABC refer to significantly different groups (Scheffé test, α=0.05), see text. / Abbildung 4: Anzahl der Nicht-Waldarten (z. B.: Schlagarten, Ruderale, Segetale und Rasen-/ Wiesenarten) in den Aufnahmegruppen Wald, großer Schlag mit Überhältern, kleine Lochhiebe, und an die Lochhiebe angrenzender Wald. *späterer Eingriff in den benachbarten Flächen! abcd und ABC beziehen sich auf signifikant unterschiedliche Gruppen (Bonferroni-Dunn test, α=0.05), siehe Text.
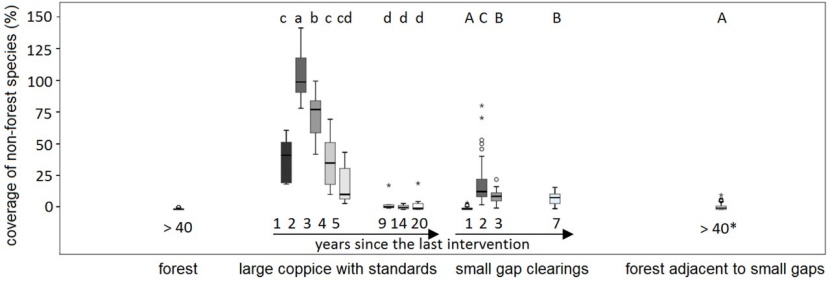
Figure 5: Cumulated coverage of non-forest species (i.e. clearcut, ruderal, segetal and turf/ meadow species) in the plot groups forest, large coppice with standards, small gap clearings, and forested plots adjacent to small gaps. * later intervention in adjacent gaps! abcd and ABC refer to significantly different groups (Scheffé test, α=0.05), see text. / Abbildung 5: Kumulierte Deckung der Nicht-Waldarten (z. B.: Schlagarten, Ruderale, Segetale und Rasen-/ Wiesenarten) in den Aufnahmegruppen Wald, großer Schlag mit Überhältern, kleine Lochhiebe, und an die Lochhiebe angrenzender Wald. *späterer Eingriff in den benachbarten Flächen! abcd und ABC beziehen sich auf signifikant unterschiedliche Gruppen (Bonferroni-Dunn test, α=0.05), siehe Text.
As demonstrated in Figure 5, non-forest species cover rises rapidly from year 1 to year 2 in the large coppice whereas in most small gaps it stays on a lower level despite their relatively high species numbers. There is no significant difference between the cover of non-forest species in the old forest and the forest plots adjacent to small gaps. Nine years after the intervention in the large coppice area, non-forest species coverage decreases to a level similar to the forest and the forested plots adjacent to the gaps, respectively. In year 20, only forest species are left in the dark environment of the newly grown canopy (Table A1): Viola mirabilis (Carpinion, L-value 4), Galium odoratum (Fagetalia, L-value 2), Polygonatum multiflorum (Querco-Fagetea, L-value 3), and Hepatica nobilis (Querco-Fagetea, L-value 4).
Carpinus betulus is by far the most frequent woody species in the tree and shrub layer (i.e. in 61 % - 100 % of relevés, Table A1). In the ground layer of the forest plots it is present in up to 60 % of relevés, in the forest plots near the gaps even in more than 60 %. Its coverage grows rapidly in the course of the years due to its ability to re-sprout (Figure A1). The increasing coverage of Carpinus reasons the strong decrease of the light-affine non-forest species.
In the large coppice with standards, Carpinion species start from a medium to low coverage, and many of them were gone or replaced by more shade tolerant ones after nine years. Further forest herbs cover by far most in the coppice with standards plots in year three. In the small gaps, many forest species raise their ground cover and keep the situation almost stable up to year seven – the last year of inventory before this part of the forest was felled. Shade tolerant Fagetalia species are less frequent in the gaps than in the forest plots. The numbers of Galio-Carpinetum and other Carpinion species in the small gaps rise from year one to year seven. Also further herbaceous species that are generally found in oak-hornbeam forests can raise their coverage remarkably until year seven (Figure 4). The light affine non-forest species remain at a level of 50 % compared to the large coppice with standards.
Discussion
Wood production has always been the main goal in forestry and thus forest management has often changed in the past decades. High forests seemed to satisfy the common demands better than coppice-with-standards forests. Not only in Central and Eastern Europe, but also in the Atlantic climate of Western Europe (Kirby et al. 2017a; Kirby et al. 2017b) a decline of regularly coppiced forests and an increase of high forest areas were observed, often at the costs of species diversity and numbers in the understorey. This decline is usually related to a darker, nutrient richer and moister environment (Hédl et al. 2010; Chudomelová et al. 2017). Nowadays, however, the conservation of near-natural ecosystems has to be taken into account, as well. Small but more frequent interventions in the forest improve the light and temperature situation for Carpinion species to an extent that may foster a more stable Carpinion community. At the beginning of the study, the forest had not been coppiced for almost 40 years (Hochbichler 2008). As a consequence, it had lost some of the characteristic Galio-Carpinetum species in the herbaceous understorey. Traditionally managed Central European oak-hornbeam coppice-with-standards forests can keep a certain degree of naturalness and therefore also a typical composition of diagnostic species (Jahn & Raben 1982; Starlinger 1997; Grabherr et al. 1998). As Becker et al. (2017) emphasized, even the proportions of plant functional types underwent a change within forty years of succession in former coppice-with- standards woodland. Species of forest edges and clearcuts had become rarer. This is in line with our findings. Only ten Carpinion species could be found, but also eight shade-tolerant Fagetalia and Querco-Fagetea species formed the herbaceous layer of the old forest about 40 years after the last coppice. In the large coppice with standards, only nine years of succession were sufficient to create a very shady situation caused by the vigorously re-sprouting hornbeam (and a few pioneer tree individuals). Hagen (2005) reports unweighted average light indicator values for oak-hornbeam communities from the same region of 5.6 and 6.2 (forest and forest edge, respectively) and 6.2 to 6.5 (clearcut). The present study shows a mean unweighted L-value of 5.2 in total, and only 4.5 for the herb layer, results that have not been shown hitherto. With respect to light conditions, Hagen´s (2005) relevés seem to be closer to traditionally managed oak-hornbeam forests. The light-indicator values of coppiced as well as non-coppiced areas are well mirrored by air temperature measurements of the present study. In the course of the 20 years without coppice, a proof of rising nutrient availability could not be found. Many species with indifferent N-values and high numbers of Fabaceae with a generally rather low demand of nitrogen may be reasons.
The large coppice leads to a strong increase in light and space and therefore fosters the enormous immigration of non-forest (especially clearcut) species immediately after the intervention. In the small gaps these species groups meet less ideal conditions. As a matter of fact, the removal of the shrub layer in the gaps also reduced the shading in the understorey of the adjacent forested plots. Nevertheless, clearcut and other non-forest species were rare, and stayed rare, throughout the years in all plots with a more or less closed tree layer. In the small gaps, more forest herbs made profit of the new light environment and less competition. In the large coppice area, clearcut species diminished gradually after a short peak period. Even forest herb numbers eventually dropped below those of the former forest. The reason is the enormous vitality of Carpinus betulus and its fast re-sprouting in the bright and warm environment. When Carpinus reached two meters in height, the understorey faced a light regime even dimmer than in the old forest. As Falinski (2003) pointed out, even periods of 10-15 years are too few to get overall insight in secondary successions and/or community stability. Data of the small gaps show that non-forest species were not able to dominate these plots, showing a tendency to decrease in number and coverage from year two to year seven. Small but more frequent interventions in the forest and the traditional use of the coppice as firewood seem to improve the light availability for Carpinion species. Thus, this method can foster a more stable Carpinion community. Yet the 20-year survey suggests that, multiple drivers have to be taken into account (Roleček et al. 2017). Very small-scaled changes in soil water content do not make things easier, especially as some of the study years were extremely dry and might have reduced the vitality of forest herbs. Even an influence of browsing animals (Laurent et al. 2017; Chudomelová et al. 2017) may count for some shifts in the herbaceous layer: Some individuals of roe deer and wild boar could obviously find a passage through - or better underneath - the fence and leave infallible signs of their presence in the study area. Thus we must ask to what extent local diversity is linked to different biogeographic, environmental, and ecosystem factors (Grace et al. 2016). Plue et al. (2017) showed in a fascinating study that only the combination of above and below ground herb species diversity (i.e. the seed bank) allows insight into the reactions of a plant community to environmental changes in broadleaved European forests. The opinion of these authors that light is one of the most important drivers in these processes supports our decision to use light as main criterion to estimate the value of different management forms. Keeping all this in mind, one should follow the conclusions of Bruckman et al. (2016) who propose a return to a soft but consequent coppice-with-standards forest management. If ever the terms “hidden” or “dark” phytodiversity (Pärtel 2014; Pärtel et al. 2016) can be applied on a managed but near-to-natural community with certainly autochthonous fluctuations as well, these proposed changes might probably bring parts of the hidden diversity back to the light.
Conclusion
Light indicator values of the herbaceous understorey react clearly and distinctly to large coppices or clearcuts, and they reflect higher daily temperatures as well as higher daily temperature fluctuations. The enormous vitality of Carpinus betulus in the study area can form a close canopy within a few years after coppicing. Non-forest species in and around small gaps never become as dominant as in largely coppiced sites. Small gaps influence the light regime (even in their direct surroundings) moderately and positively compared to the unthinned stand. More light-loving herbaceous species can re-appear even around small gaps, probably with the help of their seed bank. Large coppices with standards enable an enormous growth of non-forest species. Small but more frequent interventions into the forest would be beneficial for biodiversity and conservation, probably also for the protection of the forest from negative environmental developments. Non-forest species on a whole can be better prevented from forming a stable soil seed bank, which would emerge after large-scale interventions. The tendency towards higher summer temperatures, more irregular precipitation, more frequent damages by pests and game, and the import of alien species suggest further studies in the area. These studies will also focus on the soil seed bank of the different plots, and thus hopefully give insight in the hidden diversity of this forest.
Acknowledgements
The authors thank the forest owner family Abensberg-Traun, who gave the permission for intensive and long-term studies on silvicultural, pedological, meteorological and ecological studies. The help of numerous students and assistants of BOKU University and their engagement in often unpaid tasks was great and has to be gratefully mentioned.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors except refunding of travel costs (1999-2000, Nationalbank).
References
Axmanová, I., Chytrý, M., Zelený, D., Li, C.-F., Vymazalová, M., Danihelka, J., Horsák, M., Kočí, M., Kubešová, S., Lososová, Z., Otýpková, Z., Tichý, L., Martynenko, V.B., Baisheva, E.'v.Z., Schuster, B. & Diekmann, M. 2012. The species richness-productivity relationship in the herb layer of European deciduous forests. Global Ecology and Biogeography 21: 657-667.
Becker, T., Spanka, J., Schröder, L. & Leuschner, C. 2017. Forty years of vegetation change in former coppice-with-standards woodlands as a result of management change and N deposition. Appl Veg Sci 20: 304-313.
Bernhardt-Römermann, M., Baeten, L., Craven, D., Frenne, P. de, Hédl, R., Lenoir, J., Bert, D., Brunet, J., Chudomelová, M., Decocq, G., Dierschke, H., Dirnböck, T., Dörfler, I., Heinken, T., Hermy, M., Hommel, P., Jaroszewicz, B., Keczyński, A., Kelly, D.L., Kirby, K.J., Kopecký, M., Macek, M., Máliš, F., Mirtl, M., Mitchell, F.J.G., Naaf, T., Newman, M., Peterken, G., Petřík, P., Schmidt, W., Standovár, T., Tóth, Z., Calster, H.V., Verstraeten, G., Vladovič, J., Vild, O., Wulf, M. & Verheyen, K. 2015. Drivers of temporal changes in temperate forest plant diversity vary across spatial scales. Global Change Biology 21: 3726-3737.
Bruckman, V.J., Terada, T., Fukuda, K., Yamamoto, H. & Hochbichler, E. 2016. Overmature periurban Quercus–Carpinus coppice forests in Austria and Japan. A comparison of carbon stocks, stand characteristics and conversion to high forest. European Journal of Forest Research 135: 857-869.
Chudomelová, M., Hédl, R., Zouhar, V. & Szabó, P. 2017. Open oakwoods facing modern threats. Will they survive the next fifty years? Biological Conservation 210: 163-173.
Chytrý, M., Pyšek, P., Wild, J., Pino, J., Maskell, L.C. & Vilà, M. 2009. European map of alien plant invasions based on the quantitative assessment across habitats. Diversity and Distributions 15: 98-107.
Ellenberg, H., Weber, H.E., Düll, R., Wirth, V. & Werner, W. 2001. Zeigerwerte von Pflanzen in Mitteleuropa. 3rd ed. Verlag Erich Goltze GmbH & Co KG, Göttingen.
Essl, F., Milasowszky, N. & Dirnböck, T. 2011. Plant invasions in temperate forests. Resistance or ephemeral phenomenon? Basic and Applied Ecology 12: 1-9.
Faliński, J.B. 2003. Long-term studies on vegetation dynamics. Some notes on concepts, fundamentals and conditions. Community Ecology 4: 107-113.
Fischer, M. A., K. Oswald, and W. Adler (eds.) 2008, Exkursionsflora für Österreich, Liechtenstein und Südtirol. Bestimmungsbuch für alle in der Republik Österreich, im Fürstentum Liechtenstein und in der Autonomen Provinz Bozen / Südtirol (Italien) wildwachsenden sowie die wichtigsten kultivierten Gefäßpflanzen. Land Oberösterreich OÖ Landesmuseen, Linz.
Frost, I. & Rydin, I. 1997. Effects of competition, grazing and cotyledon nutrient supply on growth of Quercus robur seedlings. Oikos 79: 53-58.
Gerhardt, P., Arnold, J.M., Hackländer, K. & Hochbichler, E. 2013. Determinants of deer impact in European forests - A systematic literature analysis. Forest Ecology and Management 310: 173-186.
Gilliam, F.S. 2007. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 57: 845-858.
Grabherr, G., Koch, G., Kirchmeir, H. & Reiter, K. 1998. Hemerobie österreichischer Waldökosysteme. (in German)(17). Wagner, Innsbruck.
Grace, J.B., Anderson, T.M., Seabloom, E.W., Borer, E.T., Adler, P.B., Harpole, W.S., Hautier, Y., Hillebrand, H., Lind, E.M., Pärtel, M., Bakker, J.D., Buckley, Y.M., Crawley, M.J., Damschen, E.I., Davies, K.F., Fay, P.A., Firn, J., Gruner, D.S., Hector, A., Knops, J.M.H., MacDougall, A.S., Melbourne, B.A., Morgan, J.W., Orrock, J.L., Prober, S.M. & Smith, M.D. 2016. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 529: 390-393.
Hagen, R. (2005). Verjüngung, Nährstoffsituation und Wildeinflüsse auf Eichenmittelwaldschlägen des Weinviertels unter besonderer Berücksichtigung von Vereschungstendenzen. (in German). PhD-Thesis, University of Natural Resources and Life Sciences, Vienna.
Hédl, R., Kopecký, M. & Komárek, J. 2010. Half a century of succession in a temperate oakwood. From species-rich community to mesic forest. Diversity and Distributions 16: 267-276.
Hochbichler, E. 2008. Fallstudien zur Struktur, Produktion und Bewirtschaftung von Mittelwäldern im Osten Österreichs (Weinviertel). (in German). Forstliche Schriftenreihe 20: 11-246.
Jahn, G. & Raben, G. 1982. Über den Einfluss der Bewirtschaftung auf Struktur und Dynamik der Wälder. (in German). In: Dierschke, H. (ed.) Struktur und Dynamik der Wälder, pp. 717–734. J. Cramer, Vaduz.
Jamrichová, E., Szabó, P., Hédl, R., Kuneš, P., Bobek, P. & Pelánková, B. 2013. Continuity and change in the vegetation of a Central European oakwood. Holocene 23: 46-56.
Karrer, G. & Kilian, W. 1990. Standorte und Waldgesellschaften im Leithagebirge Revier Sommerein. (in German). Mitteilungen der Forstlichen Bundesversuchsanstalt: 1-244.
Kirby, K.J., Buckley, G.P. & Mills, J. 2017a. Biodiversity implications of coppice decline, transformations to high forest and coppice restoration in British woodland. Folia Geobot 52: 5-13.
Kirby, K.J., Goldberg, E.A. & Orchard, N. 2017b. Long-term changes in the flora of oak forests and of oak. Spruce mixtures following removal of conifers. Forestry 90: 136-147.
Kleinbauer, I., Dullinger, S., Peterseil, J. & Essl, F. 2010. Climate change might drive the invasive tree Robinia pseudacacia into nature reserves and endangered habitats. Biological Conservation 143: 382-390.
Laska, G. 1997. The formation of the secondary forest communities on the areas used for agricultural purposes. (in Polish, English abstract). Przeglad Przyrodnniczy 8: 77-86.
Laurent, L., Mårell, A., Balandier, P., Holveck, H. & Saïd, S. 2017. Understory vegetation dynamics and tree regeneration as affected by deer herbivory in temperate hardwood forests. iForest 10: 837-844.
Londo, G. 1976. The decimal scale for releves of permanent quadrats. Vegetatio 33: 61-64.
Oberdorfer, E. 2001. Pflanzensoziologische Exkursionsflora für Deutschland und angrenzende Gebiete. 8th ed. Ulmer, Stuttgart (Hohenheim).
Pärtel, M. 2014. Community ecology of absent species. Hidden and dark diversity. Journal of Vegetation Science 25: 1154-1159.
Pärtel, M., Bennett, J.A. & Zobel, M. 2016. Macroecology of biodiversity. Disentangling local and regional effects. New Phytologist 211: 404-410.
Plue, J., Frenne, P. de, Acharya, K., Brunet, J., Chabrerie, O., Decocq, G., Diekmann, M., Graae, B.J., Heinken, T., Hermy, M., Kolb, A., Lemke, I., Liira, J., Naaf, T., Verheyen, K., Wulf, M., Cousins, S.A.O. & Cáceres, M. de 2017. Where does the community start, and where does it end? Including the seed bank to reassess forest herb layer responses to the environment. Journal of Vegetation Science 28: 424-435.
Pyšek, P. & Chytrý, M. 2014. Habitat invasion research. Where vegetation science and invasion ecology meet. Journal of Vegetation Science 25: 1181-1187.
Roleček, J., Vild, O., Sladký, J. & Řepka, R. 2017. Habitat requirements of endangered species in a former coppice of high conservation value. Folia Geobot 52: 59-69.
Schume, H. & Starlinger, F. 1996. Boden- und vegetationskundliche Gliederung von eichenreichen Wäldern im östlichen Österreich. (in German). FBVA-Berichte: 11-60.
Starlinger, F. 1997. Natürliche Waldgesellschaften im "Sommerwarmen Osten" Österreichs. (in german). FBVA-Berichte 95: 45-58.
Ulrich, E. (1987). Bodenwasserhaushalt eines Eichenmittelwaldstandortes (mit Mistelbefall). Diplomarbeit, University of Natural Resources and Life Sciences, Vienna.
Willner, W., and G. Grabherr (eds.) 2007, Die Wälder und Gebüsche Österreichs. Ein Bestimmungswerk mit Tabellen (in German). Elsevier Spektrum Akad. Verl., München.
Appendix
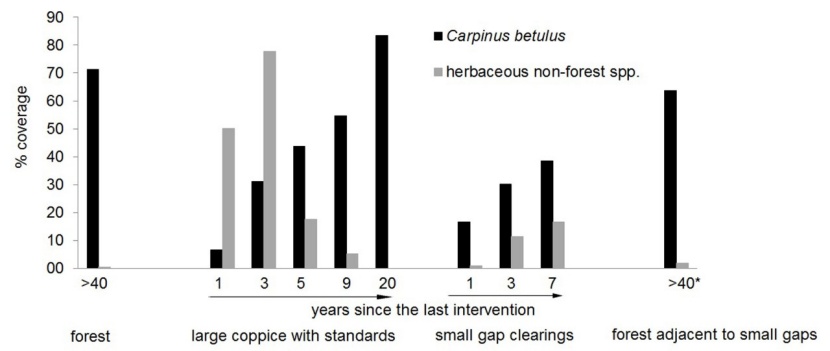
Figure A1: Average coverage of Carpinus betulus in comparison with herbaceous non-forest species in the plot groups forest, large coppice with standards, small gap clearings, and forested plots adjacent to small gaps. * later intervention in adjacent gaps! / Abbildung A1: Mittlere Deckung von Carpinus betulus in Vergleich mit krautigen Nicht-Waldarten in den Aufnahmeflächen Wald, großer Schlag mit Überhältern, Lochhiebe und Wald angrenzend an die Lochhiebe. *späterer Eingriff in angrenzenden Lochhiebe.
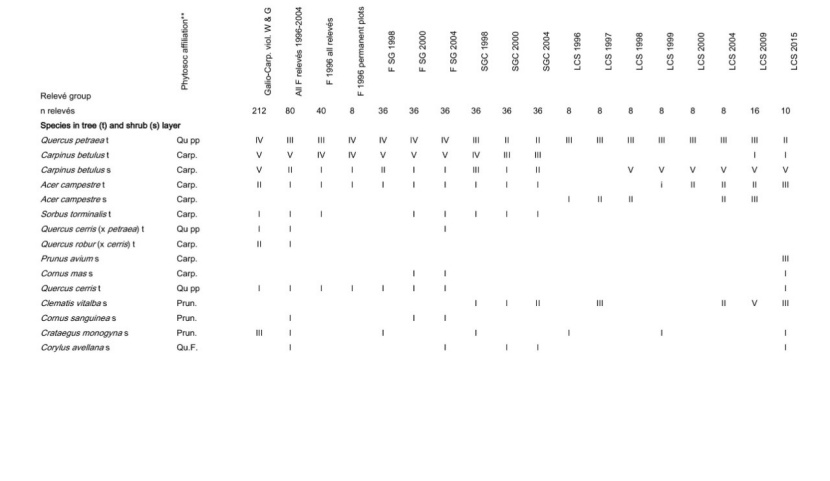
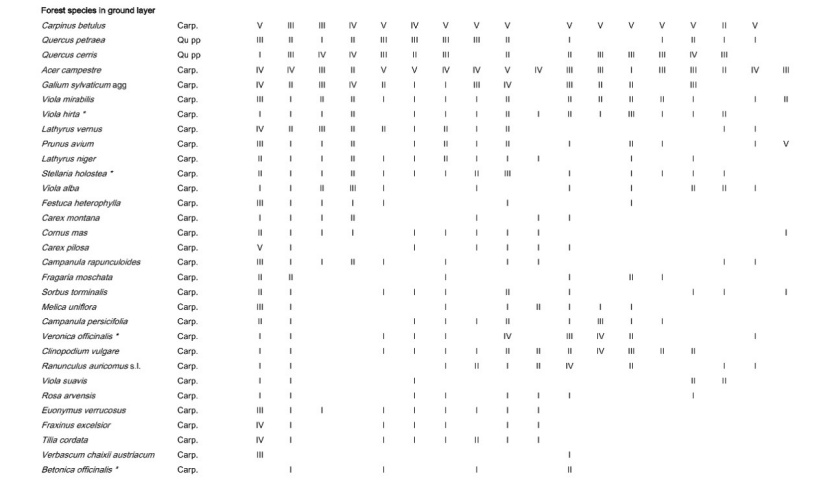
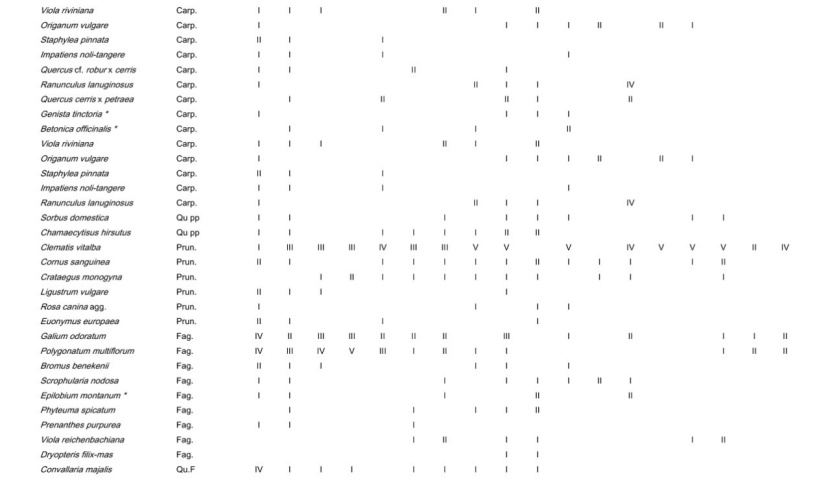
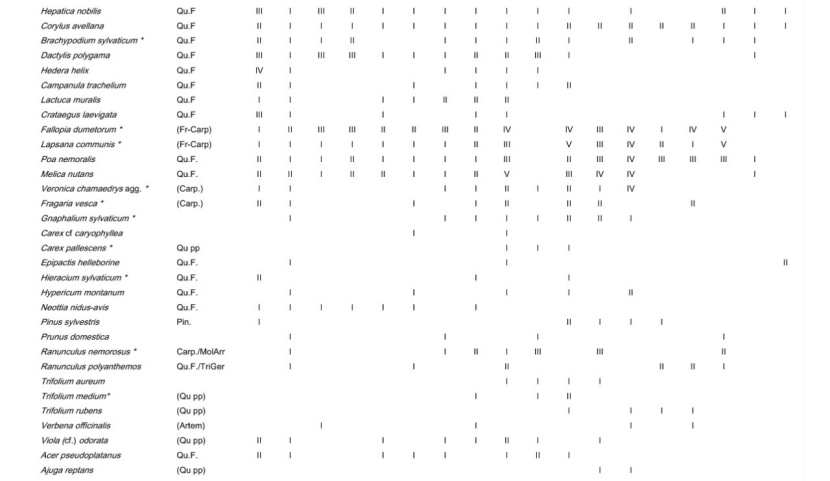
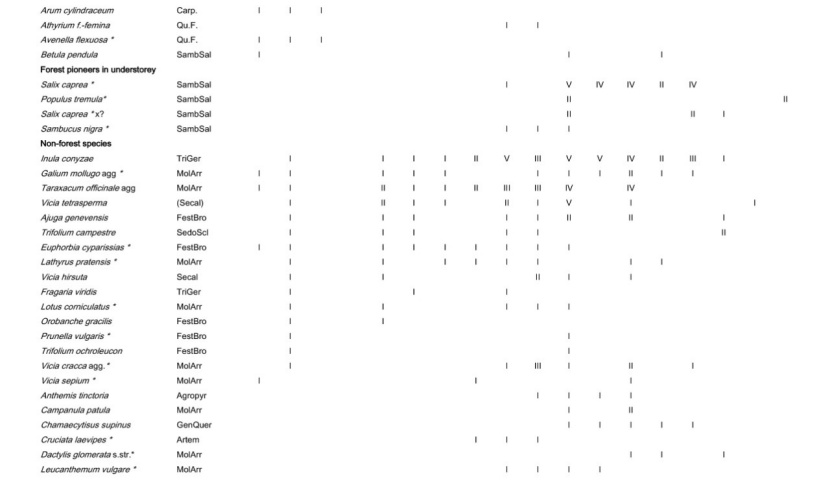
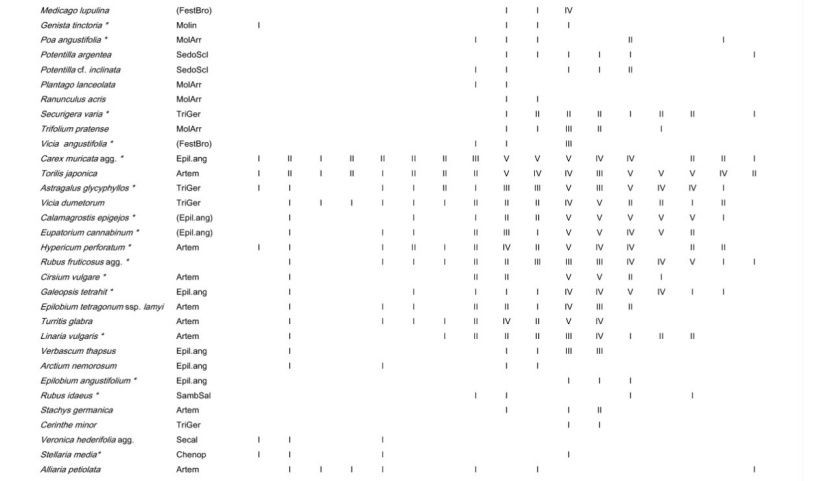
Table A1: Frequency of all species in the different plot groups, F – forest, FSG – forest adjacent to small gaps, SGC – small gap clearings, LCS –large coppice with standards, 1st column indicates the frequency (I: 1-20 % of the relevés contain the species II: 21-40 % a.s.o. V: 81-100 %) in Galio sylvatici-Carpinetum violetosum mirabilis published in Willner and Grabherr, 2007.
Tabelle A1: Stetigkeit sämtlicher Arten in den verschiedenen Aufnahmegruppen, F-Wald, FSG-Wald angrenzend an Lochhiebe, SGC-Lochhiebe, LCS-großer Schlag mit Überhältern, 1ste Spalte gibt die Stetigkeit (I: 1-20 % der Aufnahmen enthalten die Art, II: 21-40 % usw. V: 81-100 %) im Galio sylvatici-Carpinetum violetosum mirabilis aus Willner und Grabherr, 2007, an.