Schlüsselbegriffe: Wiederaufforstung, Hyrkanische Wälder, Streu, Klimawandel, Sonnenstrahlung, Mitigation
Abstract
Albedo and carbon sequestration, as important climatic parameters, are influenced by many factors. Since forests are important factors affecting carbon stocks, albedo, they have a significant role in global warming. This study intends to explore interactions between trees species and albedo, soil carbon (C) and nitrogen (N) stocks in the Hyrcanian forest of northern Iran. The study sites consist of five 20-year-old forest plantations consisting of maple (Acer velutinum Bioss), poplar (Populus deltoides Barter.ex Marsh), Mediterranean cypress (Cupressus sempervirens var. Horizontalis (Mill.) Loudon), chestnut-leaved oak (Quercus castaneifolia C. A. M. macranthera F. & M.) and alder (Alnus glutinosa (L.) Gaertn). In the summer, ten randomly located soil samples were taken from each stand from the top 10 cm of the soil to determine the soil characteristics, soil organic C, total C and N. Albedo was estimated with an albedometer set-up, which was installed on a movable mast in each stand. We used two analytical methods to determine the effect of tree species on albedo and soil C and N stocks. ANOVA was used to determine the significant difference between tree species and albedo, soil C and N stocks. TOPSIS technique – a commonly used classical Multi-Criteria Decision Making (MCDM) method – was employed to prioritize tree species considering the combination of the examined factors (albedo, C and N stocks). Our analysis suggests significant differences in albedo and soil C and N stocks between tree species (p < 0.01). Albedo of the alder stand (0.33) was highest and for Mediterranean cypress it was lowest (0.15). The highest values of soil C and N stocks we observed in the alder stand (64.61 and 2.68 t ha-1, respectively) and the lowest amounts in the poplar stand (26.76 and 1.2 t ha-1 respectively). The results of TOPSIS ranking indicated that, the preferred tree species are alder, chestnut-leaved oak, maple, polar and Mediterranean cypress, respectively. The findings of this study demonstrated that different planted tree species have different substantial effects on albedo and soil properties in the study region. Our results demonstrate the benefits of intercropped broad-leaved trees on albedo and soil C and N stocks. These findings may help to optimize forest management activities, such as selection of tree species in reforestations, plantations and agroforestry systems.
Zusammenfassung
Albedo und Kohlenstoffspeicherung sind wichtige klimatische Parameter und werden von mehreren Faktoren beeinflusst. Baumarten sind einer dieses Einflussfaktoren und haben letztendlich auch Auswirkung auf die globale Erwärmung. Diese Studie unersucht den Effekt verschiedener Baumarten auf Albedo- und Bodenkohlenstoff (C) und Stickstoff (N) Speicherung. Unser Untersuchungsgebiet ist eine 20 Jahre alte Aufforstung mit Ahorn, Erle, Eiche, Zypresse und Pappel im Hyrkanischen Wald im Norden des Iran. Im Sommer wurden in jedem untersuchten Bestand jeweils zehn Proben zufällig aus den ersten 10 cm der obersten Bodenschicht entnommen um die Bodeneigenschaften zu untersuchen, insbesondere Bodentextur, Feuchtigkeit, organischem C, Gesamt-C und N. Die Albedo wurde mit einem Albedometer gemessen, das an jedem Standort auf einem beweglichen Mast installiert wurde. Wir verwendeten hier zwei Analysemethoden, um die Bedeutung von Baumarten auf Albedo und Boden C und N Vorrat abzuschätzen. Mit ANOVA wurden signifikante Unterschiede in Albedo und Kohlenstoff- und Stickstoffbindung zwischen den Baumarten geprüft. Die TOPSIS-Technik – eine oft verwendete klassische MCDM-Methoden (Multi-Criteria Decision Making) – erlaubte uns die Priorisierung der Baumarten unter Berücksichtigung der Kombination der drei untersuchten Faktoren zu ermitteln. Unsere Analysen zeigten signifikante Unterschiede zwischen Albedo, Boden C und N Speicherung zwischen den Baumarten (p < 0.01). Die Albedo im Erlenbestand (0.33) war am höchsten und in dem Zypressenbestand am niedrigsten (0.15). Die höchsten Werte für die Kohlenstoff- und Stickstoffspeicherung im Boden wurden im Erlenbestand beobachtet (64.61 bzw. 2.68 t ha-1) und die niedrigsten Mengen im Pappelbestand (26.76 bzw. 1.2 t ha-1) erfasst. Die Ergebnisse des TOPSIS-Rankings zeigten folgende optimale Reihenfolge der untersuchten Baumarten: Erle, Eiche, Ahorn, Pappel und Zypresse. Die Ergebnisse dieser Studie zeigten, dass verschiedene Baumarten unterschiedliche signifikante Auswirkungen auf die Albedo und Bodeneigenschaften in der Untersuchungsregion haben. Wir konnten zeigten, dass insbesondere Laubbäume vorteilhafte Albedo und Boden C und N Speicherung aufweisen. Diese Erkenntnisse können dazu beitragen, die Waldbewirtschaftung zu optimieren, wie etwa die Auswahl von Baumarten für Wiederaufforstungen, Plantagen und Agroforestry-Systemen.
1. Introduction
Climate change is one of the most important drivers for land management, since it has adverse effects on the environment (Nik et al., 2015). Forests play a significant role in mitigating the climate change impacts (e.g., Sohngen and Mendelsohn, 2003; Kindermann et al., 2008) and cooling the Earth’s surface in tropical and temperate regions (Ellison et al., 2017). Considering the importance of forests to mitigate the consequences of climate change, protection of forests and adopting strategy to increase the forested area, are urgently needed (Ellison et al., 2017). Forest development is the one of the most important economic and ecological solution for contrasting climate change (Kuusinen et al., 2016; Scheidel, 2016; Streck, 2012; Ellison et al., 2017; Templer et al., 2012). Forest managers have extended forests by plantation, reforestation and afforestation (Angelsen and Kanounnikoff 2008).
Recent studies have investigated the role of afforestation on climate change through its impact on the forest albedo and carbon stocks (e.g. Bala et al. 2007). Trees influence local climate via carbon sequestration (Bonan, 2008; Smith et al., 2012), changing surface albedo (Myhre et al., 2013) and affecting greenhouse gas emissions (Canadell and Raupach 2008; Ellison et al., 2017).
Albedo is one of the most important and effective factors on climate (Lutz and Howarth, 2014). Albedo is the ratio of incoming solar radiation to the reflection of the Earth’s surface (Burakowski et al., 2015). The importance of albedo implications of land use change and land management change on climate change mitigation is widely acknowledged in the scientific community (Betts, 2000; Bright et al., 2015; Davies-Barnard et al., 2015). High albedo values cool the earth surface and the magnitude of cooling depends on variety of factors including snow depth, cloud cover, solar zenith angle, land cover type, growth conditions and soil moisture (Mote, 2008; Atlaskina et al., 2015). Forest species composition and canopy structure affect the reflected radiation and in turn affect the climate (Stephens et al., 2015; Alkama and Cescatti, 2016; Naudts et al., 2016; Hovi et al., 2016). Differences in reflection among tree species are linked to differences in foliage, structure of canopies and the proportion of trees (Roberts et al., 2004; Rautiainen and Stenberg, 2005; Smolander and Stenberg, 2005; Kuusinen, 2014). Coniferous trees have, in general, lower reflection than broad-leaved trees (Kuusinen et al., 2014; Hollinger et al., 2010). In mixed conifer-broadleaved forests, albedo was found to be linearly increasing with increasing proportion of deciduous trees (Lukeš et al., 2013a).
Another factor affecting the climate is carbon (C) sequestration. Forest ecosystems play a major role in the exchange of CO2 between atmosphere and biosphere (Sariyildiz, 2015; Kaul, 2010; Templer et al., 2012). Approximately 75 % of terrestrial C content is stored in the soils (Goodale et al., 2002). Forests store nearly 45 % of terrestrial C and about two-thirds of such amount is retained in the soil (Goodale et al., 2002). In fact, C stock inside soils is one of the most important ecosystem services because of its role in climate regulation (IPCC, 2007). Forest trees and soils are the main sinks to absorb atmospheric gases (Kaul, 2010; Templer et al., 2012). Quantifying forest soil C stock is critical to understand the ecological responses of forests to climate change, land use management and to improve global change models (Smith et al., 2012; Dib et al., 2014). C stock in forest soils (Lal, 2005) is affected by natural and anthropogenic factors (Larionova et al., 2002) such as tree species, soil texture, land use pattern and soil moisture (Jana et al., 2009).
Another important soil element is nitrogen (N). Soil N content is an important parameter for assessment of tree species effects on ecosystem functioning. Recently, N stocks have been recognized as important indicators of C sequestration capacity in soils (Akselsson et al., 2005; De Vries et al., 2006). The variability in soil C and N stocks may be linked with tree species composition, differences in the thickness of the forest floor, microbial processes and the quality and quantity of tree species (Sariyildiz et al., 2015; Osher et al., 2003; Marcos et al., 2010; Smolander and Kitunen, 2002; Menyailo et al., 2002; Lovett et al., 2002).
Variation of the amount of forest floor and the biochemical properties of coniferous and deciduous tree species affects forest floor decomposition and influence soil C stocks (Sariyildiz and Anderson, 2005). For example, forest floor of coniferous tree species will usually decay relatively slow and create a high C stock in the forest floor over time (Finzi et al., 1998; Lovett et al., 2004; Vesterdal et al., 2008). Vesterdal et al., (2008) indicated that tree species influence the stocks and composition of soil C, which may also affect other soil characteristics (Marcos et al., 2010) and/or alter the potential of the soil to supply nutrients to tree species (Prescott and Vesterdal 2013; Aponte et al., 2013; Yatso and Lilleskov, 2016).
Hyrcanian forests in the North of Iran are among the oldest forests in the Northern hemisphere with unique richness of biological diversity, endemic tree species and various vegetation landscape (Akhani et al., 2010). Nowadays, the area of these forests have been degraded (Kooch et al., 2014), consequently reforestation and plantation with various tree species are needed for rehabilitation of these degraded forests. Therefore, selection of the most suitable tree species is very important to optimize forest regeneration operations under global warming.
The objective of this study is to determine the suitable tree species for forest development under global warming conditions considering the combination of albedo and C and N stocks. We tested the following hypotheses: do broad-leaved tree species have higher albedo, soil C and N stocks than coniferous trees?
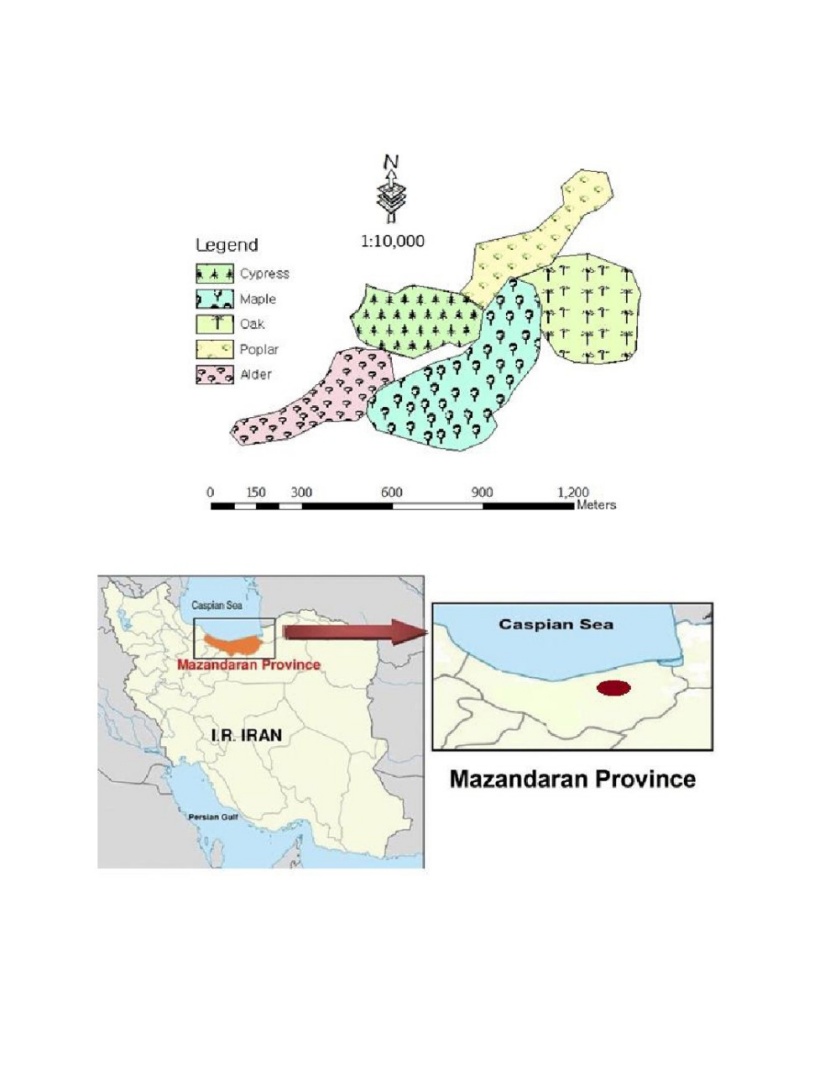
Figure 1: Site study in Hyrcanian forests (North of Iran). / Abbildung 1: Untersuchungsgebiet in Hyrkanischen Wäldern (Nordiran).
2.1 Study area and field plots
The study area is situated in the North of Iran (36°,07‘,20‘‘ to 36°,22‘,25‘‘ N latitudes and 53°,02‘,25‘‘ to 53°,07‘,20‘‘ E longitudes) (Figure. 1). The study sites are located between 230 and 250 m a.s.l. with similar climatic conditions. The minimum and maximum temperatures were recorded in January (6.4˚C) and August (27.1˚C), respectively. Annual mean rainfall is 923 mm (Mahdasht meteorological station). The climate is temperate-humid. According to the USDA Soil Taxonomy, soils can be classified as sandy-clay-loam to clay-loam Alfisols, developed on marny limestones originating from Cretaceous to Jurassic periods (Anonymous, 2011; Anonymous, 2015). Previously, these areas were dominated by natural forests dominant by chestnut-leaved oak (Quercus castaneifolia C. A. M. macranthera F. & M.), hornbeam (Carpinus betulus L.) and ironwood trees (Parrotia persica (DC) C.A.Mey). In the 1990s, these forests were clear-cut. In some areas after clear cutting, reforestation has been done using a spacing of 2 × 2 m with various tree species including maple (Acer velutinum Bioss), poplar (Populus deltoides Barter.ex Marsh), Mediterranean cypress (Cupressus sempervirens var. Horizontalis (Mill.) Loudon) (from now on we use Cypress instead of Mediterranean cypress to be more concise in the text), chestnut-leaved oak (Quercus castaneifolia C.A.M. macranthera F. & M.) (from now on we use Oak instead of Chestnut-leaved oak to be more concise in the text) and alder (Alnus glutinosa (L.) Gaertn) (Figure 1). Three hectares area of each planted forest were studied to examine the effect of forest tree species on canopy albedo and soil properties. The planted stands were never fertilized and had the same forest management system. Thus, in a relatively small homogeneous area, reforested stands with similar conditions can be found (Anonymous, 2015; Anonymous, 2011).
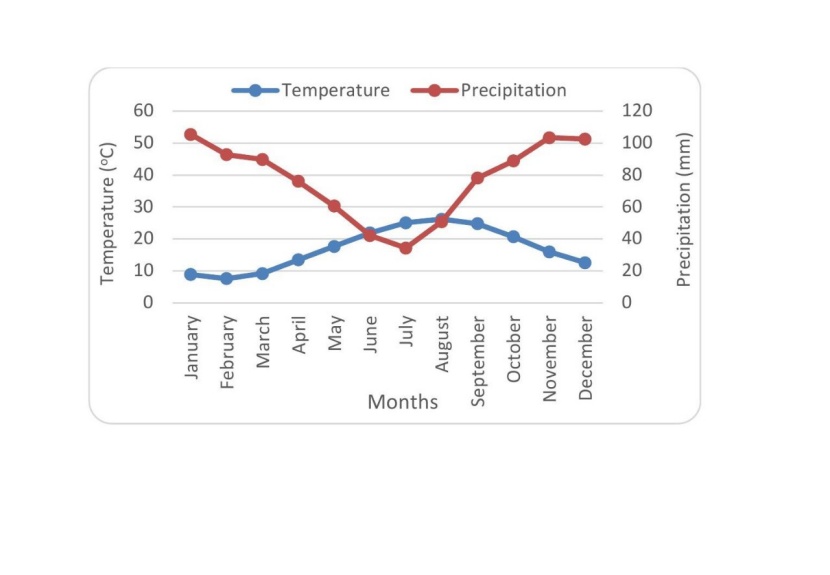
Figure 2: Mean monthly temperature and precipitation in study area based on Mahdasht meteorological station data. / Abbildung 2: Mittlere monatliche Temperatur und Niederschlag im Untersuchungsgebiet, basierend auf Daten der Mahdasht-Messstationen.
Field sampling
Ten sample plots of 20×20 m were placed within each of the five stands. All of the sample plots in each site had similar slope, elevation (230 - 250 m a.s.l) and aspect (north). Trees at the edge of the stands were not considered during sampling, to eliminate border effects. Measurements were made between July and August 2018. Total tree height, DBH diameter at breast height), stand density (number of live tree per hectare), crown length and diameter, percentage of living crown, slope and elevation of study sites were measured in each plot (Table.1).
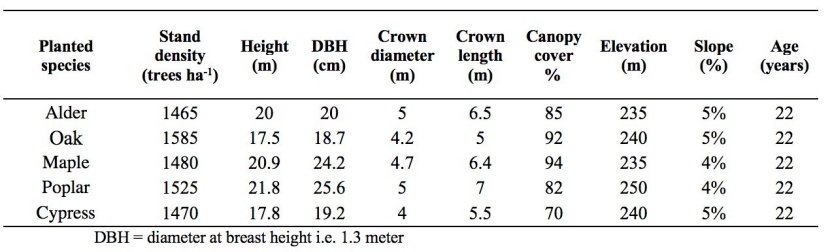
Table 1: Stand variables average (Acer velutinum Bioss, Populus deltoides Barter.ex Marsh, Cupressus sempervirens var. Horizontalis (Mill.) Loudon), Quercus castaneifolia C. A. M. macranthera F. & M.) and Alnus glutinosa (L.) Gaertn. / Tabelle 1: Mittelwert der Standortvariablen (Acer velutinum Bioss, Populus-Deltoides Barter.ex Marsh, Cupressus sempervirens var. Horizontalis (Mill.) Loudon), Quercus castaneifolia C. A. M. macranthera F. & M.) und Alnus glutinosa (L.) Gaertn.
2.2 Methods
In-situ albedo measurement
Forest albedo was measured from the ground level measurements, which used the direct normal radiation and the Earth’s reflection using a portable albedo-meter. The used instrument was a CM-7B manufactured and calibrated by Kipp & Zonen, which measures the albedo in the wavelengths ranging from 300 to 3000 nm (Hollinger et al., 2010; Thiel et al., 2008).
The albedo data were collected during the summer from July to August in each stand. The albedo-meter was positioned horizontally about 2 m above the canopy and thus captured the entire forest tree cover. Nearly all of the measurement data were taken during full sunny days. The daily albedo were measured over 24 hours. Albedo was calculated using the ratio of daily total reflection (W m-2) to the incident solar radiation (W m-2) (Equation 1) (Kuusinen 2014).
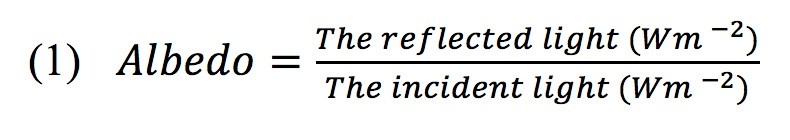
Mineral soil and forest floor sampling and analysis
Soil sampling was carried out during the summer in August. To take the soil samples, the organic soil layer was removed over the mineral soil. Ten soil profiles (40×40cm) were dug in each stand (10 replicates), resulting in 50 mineral soil samples (i.e., 10 plots at each site × 1 soil sample in the center of each plot × 5 sites) at a depth of 0–10cm (without L and FH layers) (LA fleur et al., 2015) and 50 forest floor samples (only L and FH layers) were taken from each stand.
The forest floor samples were dried and grounded. Total C and N contents were determined using dry combustion with an elemental analyzer (Fisons EA1108, Milan, Italy) calibrated by the BBOT standard (Thermo Quest Italia S.p.A.). The mineral soil samples were air-dried and passed through a 2-mm sieve. Soil water content was then measured following drying the samples in an oven at 105 °C for 24 h. Bulk density was measured by the Clod method (Blake and Hartge, 1986; Akin and Özdemir, 2003). The soil bulk densities is measured by the ratio of mass and volume. Soil texture was determined by the Bouyoucos hydrometer method (Bouyoucos 1962). Soil pH was detected using an Orion Analyzer Model 901 pH meter in a 1:2.5 soil:water solution and soil EC (electrical conductivity) were determined using an Orion Ionalyzer Model 901 EC meter in a 1:2.5 soil:water solution (Parsapour et al., 2018). Soil organic carbon was determined by Walkley-Black technique (Allison, 1975) and total nitrogen was measured based on the semi Micro-Kjeldahl technique (Bremner and Mulvaney, 1982).
Soil C and N stocks were calculated using the following equations (Sariyildiz et al., 2015):
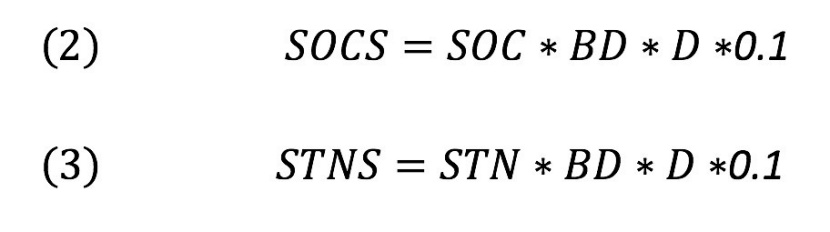
With BD being the bulk density (g cm-3), SOC soil organic carbon (g kg-1), STN soil organic nitrogen (g kg-1), D soil thickness (cm), SOCS soil organic carbon stocks (t ha-1), STNS soil organic nitrogen stocks (t ha-1).
2.3 Statistical analysis
Normality of the variables was checked by Kolmogorov-Smirnov test and Levene’s test was used to examine the equality of the variances. One-way analysis of variance (ANOVA) was used to compare the significant difference in forest tree albedo values and soil C & N stocks among various stands. SNK (Student-Newman-Keuls) test was employed for grouping of tree species considering one of these factors (Albedo values or soil C & N stocks) at p < 0.05 level. All statistical analyses were conducted using the SPSS 24.0 statistical software package. Linear regression analysis was used to test for correlations among soil properties and albedo among plantation forests.
In order to prioritize the most appropriate species for climate change mitigation efforts in Northern Iran, TOPSIS technique was utilized (Using the Excel software). The TOPSIS technique was used to integrate all the research data into a final ranking by considering the results of all the related factors (albedo, soil C and N stocks). TOPSIS (technique for order performance by similarity to an ideal solution) is one of the famous and classical Multi-Criteria Decision Making (MCDM) methods, which was initiated for the first time by Hwang and Yoon (1981). The basic concept of this method is that the best selected alternative should simultaneously minimize the distance from an ideal point and maximize the space from a nadir point in a geometrical sense (Olson, 2004). TOPSIS algorithm determines the most preferable choice among all the possible choices. The best alternative is the one with the shortest distance to the fuzzy positive ideal solution and with the longest distance to the fuzzy negative ideal solution. There have been some studies in the literature using TOPSIS for the solution of MCDM problems (Olson, 2004; Uzun and Yıldırım, 2016). The TOPSIS procedure is shown in Figure 3 in five main steps (Shyur et al., 2006):

Figure 3: Procedure of TOPSIS method. / Abbildung 3: Die Vorgehensweise der TOPSIS-Methode.
3. Results
3.1 Albedo
Our results indicated that there were significant differences in albedo among tree species. Albedo changed by various tree species. Daily albedo amounts varied between 0.157 (Cypress) and 0.338 (Alder) (Figure.4). Mean albedo was found in the ranked order of alder > poplar > maple > oak > cypress. Albedo in the alder (0.33) was higher (p < 0.01) than in the other stands (Figure. 5). Cypress had the lowest summer albedo. The establishment of plantations with coniferous and broad-leaved trees resulted in changing albedo with broad-leaved trees exhibiting significantly higher albedo than coniferous tree species. In this study, the amount of albedo in the broad-leaved trees was 62-76 % higher than that in the coniferous trees. The low albedo of cypress trees means that they absorb a large share of the incoming solar radiation and are more strongly heated up as broad-leaved trees. In turn, coniferous species may warm up the surrounding air more intensively. Oak and maple of our study sites had equal amounts of albedo, which was lower than alder and poplar trees.
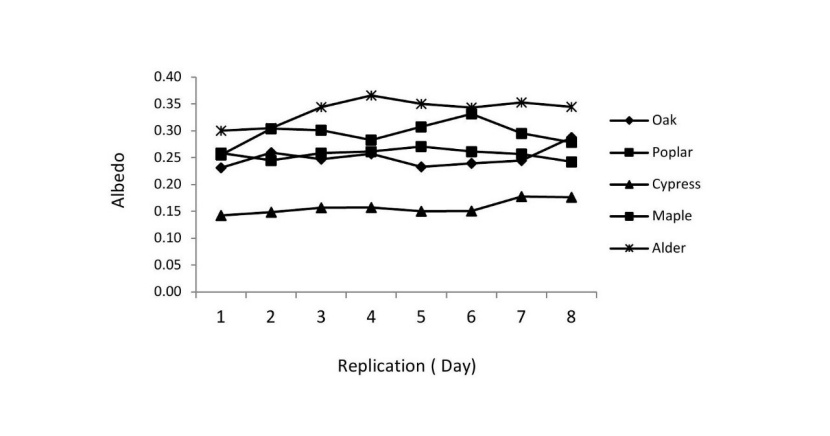
Figure 4: Daily albedo in summer of the five studied stands. / Abbildung 4: Tägliche Albedo im Sommer der fünf untersuchten Beständen.
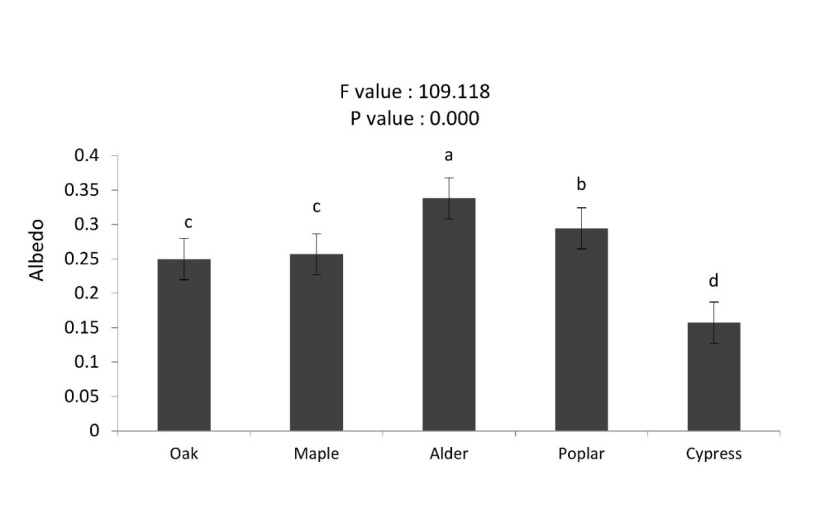
Figure 5: Mean (±SE; n = 8) of summer albedo in different tree species. / Abbildung 5: Mittelwert (± SE; n = 8) der Sommeralbedo in verschiedenen Baumarten.
3.2 Forest floor and soil properties
All forest floor properties showed statistically significant differences among different tree species. Carbon concentrations of forest floor were significantly higher in the poplar, maple and alder compared to oak and cypress. Nitrogen concentrations of forest floor were significantly different among tree species (alder ≈ maple > oak ≈ cypress≈ poplar, Figures 6 and 7).
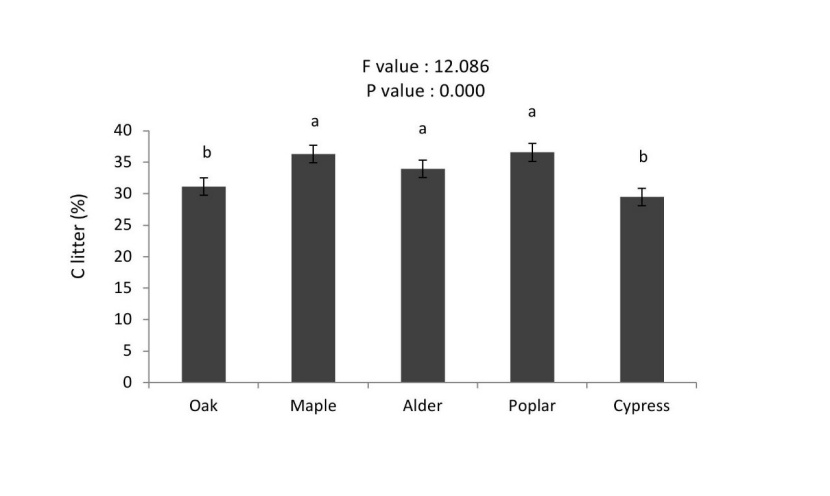
Figure 6: Mean (±SE; n = 10) of forest floor carbon (%) in different tree species. / Abbildung 6: Mittelwert (± SE; n = 10) von Kohlenstoff in der Streuschicht (%) in verschiedenen Baumarten.
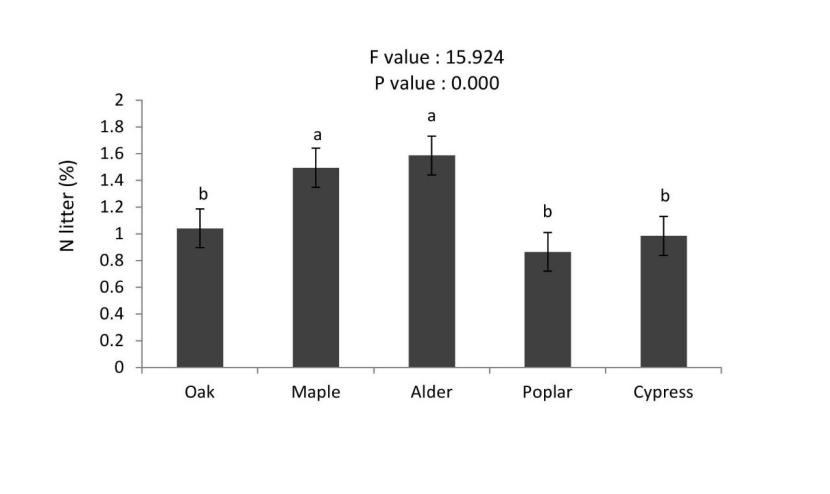
Figure 7: Mean (±SE; n = 10) of forest floor N in different tree species. / Abbildung 7: Mittelwert (± SE; n = 10) der Streu N in in verschiedenen Baumarten.
3.3 Soil carbon and nitrogen stocks
Soil properties are presented in table 2. Our data showed that there were significant differences in the soil features among different land covers (Table. 2).
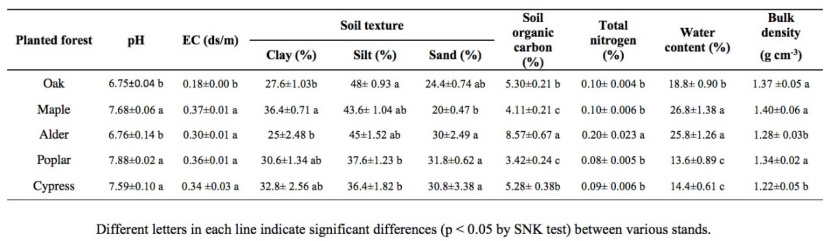
Table 2: Stand variables description of the five examined stands. / Tabelle 2: Boden der Standortsvariablen der fünf untersuchten Bestände.
Soil carbon stocks were significantly higher in alder stand than oak > cypress > maple > poplar stands (Figure 8). Total soil nitrogen stocks in the alder was higher than other stands (2.68 t N ha-1), whereas no significant differences with poplar, maple, cypress and oak stands were detected (Figure 9). The soil organic carbon (SOC) was significantly higher under plantations of alder > oak ≈ cypress > maple ≈ poplar stands (Figure 10).
C/N ratios under cypress ≈ oak were significantly higher than under alder ≈ maple ≈ poplar stands (Figure 11). The soil C/N ratio differed significantly among tree species (p < 0.01). The cypress stand had the highest C/N ratio and poplar the lowest C/N ratio among the examined tree stands.
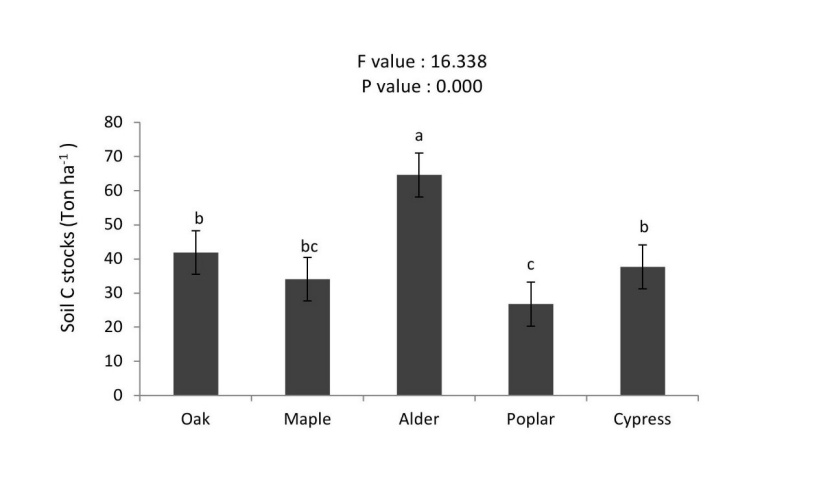
Figure 8: Mean (±SE; n = 10) of soil carbon stocks under different tree species. / Abbildung 8: Mittelwert (± SE; n = 10) der Bodenkohlenstoffbindung unter verschiedenen Baumarten
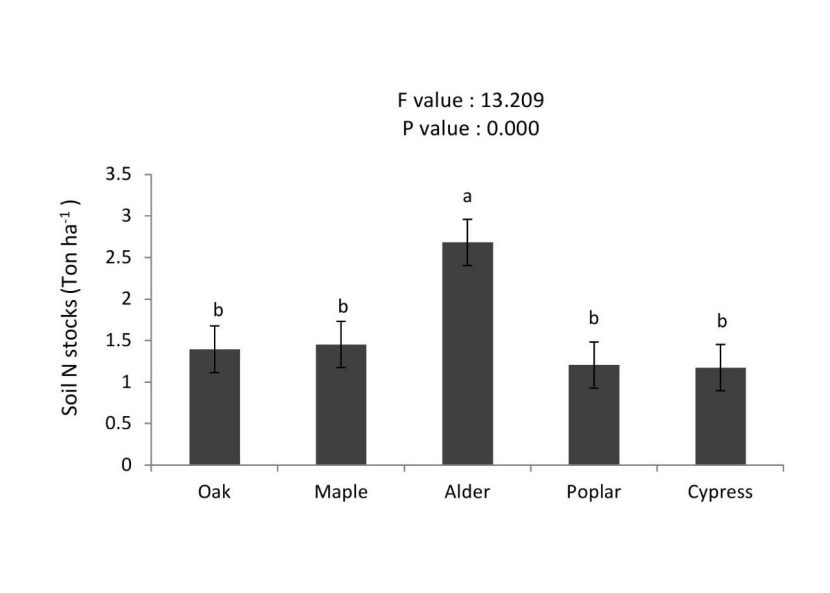
Figure 9: Mean (±SE; n = 10) of soil nitrogen stocks under different tree species. / Abbildung 9: Mittelwert (± SE; n = 10) der Bodenstickstoffbindung unter verschiedenen Baumarten.
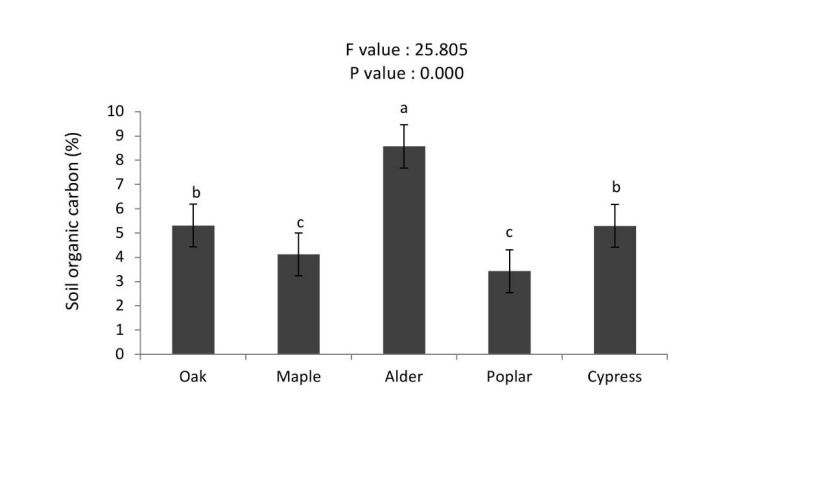
Figure 10: Mean (±SE; n = 10) of soil organic carbon in different tree stands. / Abbildung 10: Mittelwert (± SE; n = 10) der organische Bodenkohlenstoff in verschiedenen Baumarten.
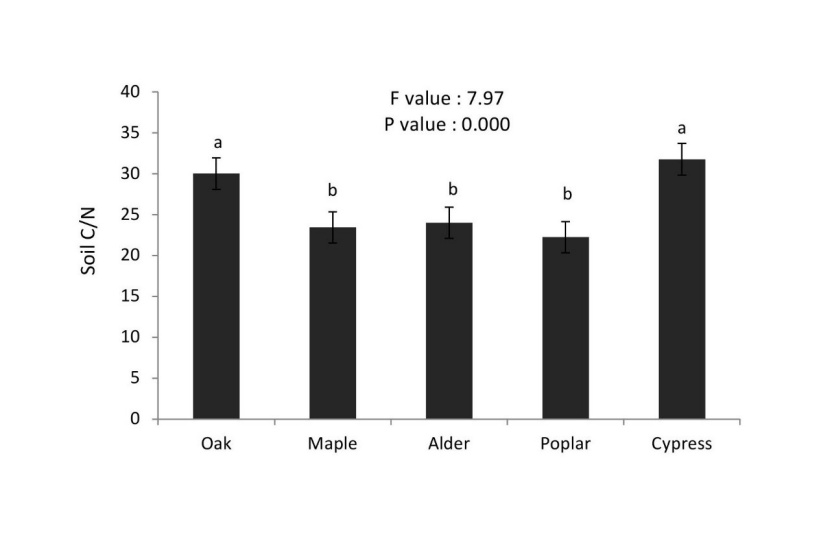
Figure 11: Mean (±SE; n = 10) of carbon/nitrogen ratio in different tree stands. / Abbildung 11: Mittelwert (± SE; n = 10) des Kohlenstoff/Stickstoff Verhältnis in verschiedenen Baumarten.
3.4 Regression analysis of variables
Regression analysis of variables showed that there are significant positive correlations between soil carbon and nitrogen stocks. Stands with higher soil carbon had also higher nitrogen stocks in the soil and the C content had a highly significant positive correlation with the N content (R2 = 0.73, p < 0.01). Albedo was correlated with the Earth reflection (R2 = 0.93, p < 0.01) at all the study sites. Soil C and N contents decreased with increasing soil clay content (Figure. 12a, b, c, d).
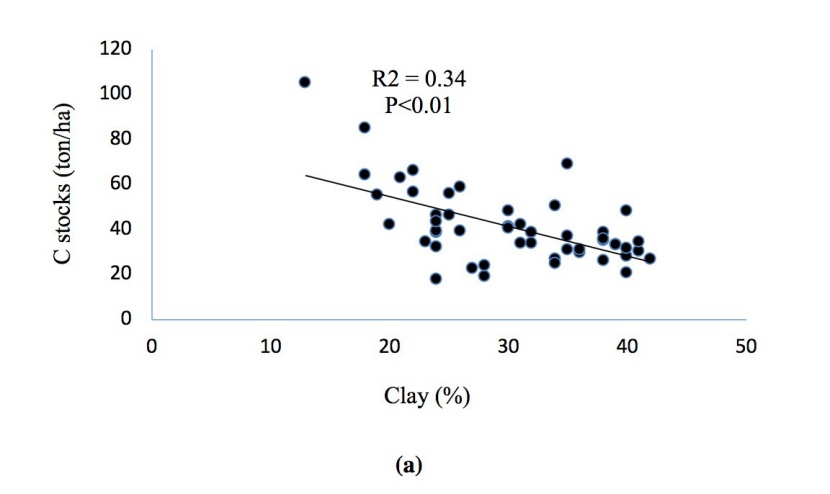
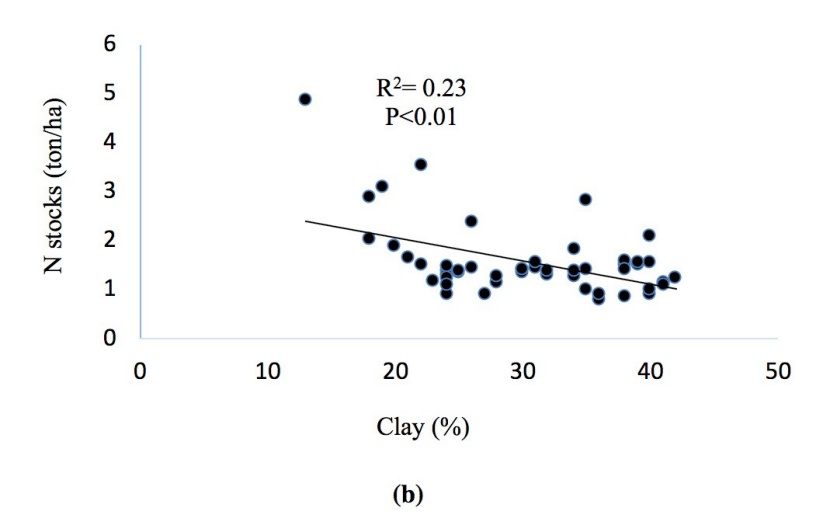
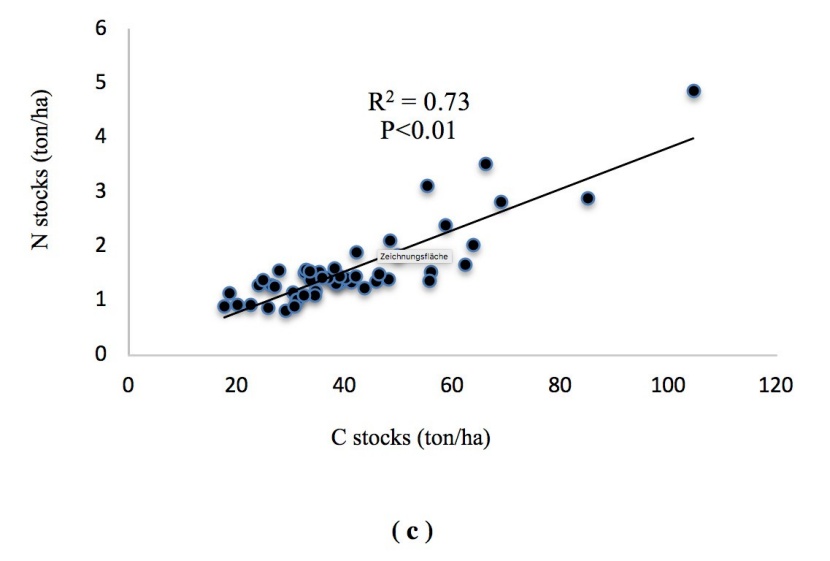
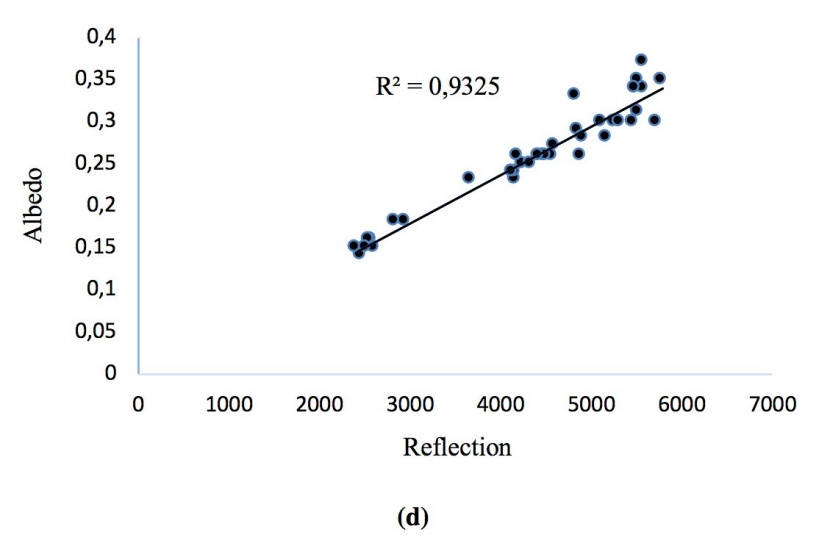
Figure 12: Correlations of (a) soil C and clay, (b) soil N and clay, (c) soil C and N stocks, (d) albedo and reflection.
Abbildung 12: Korrelationen von (a) soil C and clay, (b) soil N and clay, (c) soil C and N stocks, (d) albedo and reflection.
3.5 Ranking of tree species based on TOPSIS technique
The results of TOPSIS technique are presented in Table 3. Alder with the maximum score in TOPSIS ranking is the first tree species suggested for forest development in the study region. Based on the TOPSIS analysis, overall ranking for the preference of species to develop in the forested areas under global warming conditions are alder, oak, maple, poplar and cypress, respectively.
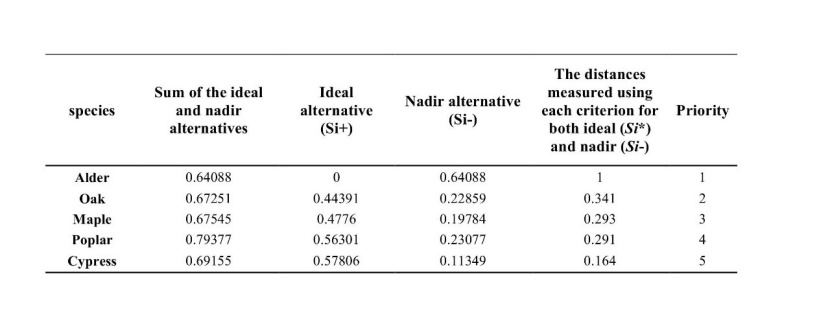
Table 3: Ranking of the optimum forest tree species considering the albedo, and carbon and nitrogen stocks. / Tabelle 3: Ranking der optimalen Waldbaumarten unter Berücksichtigung der Albedo, Kohlenstoff und Stickstoff-Speicherung.
4. Discussion
Our detailed assessment of tree species effects on the albedo and the soil C and N stocks in the forest plantation in Northern Iran show that these factors are affected by tree species and the magnitude of the effect depended on a number of factors, most importantly, land-cover and tree species (Lara-Romero et al., 2017).
4.1 Albedo
Our results indicate that there is a complex interaction between tree species and albedo. Albedo varied between broad-leaved or coniferous species. The variation within the broad-leaved tree species was also large. Our results showed that broad-leaf tree species in this region had higher albedo (higher than 0.2) than conifers (lower than 0.2). These results coincide with previous literature suggesting that albedo mainly changed according to land cover type, tree species composition and canopy structure in forests (Atlaskina et al., 2015; Kuusinen et al., 2014, 2016; Lutz and Howarth 2014; Lukeš et al., 2013b). Coniferous needles tend to have lower albedo than the leaves of broad-leaved trees, similar to the results of Kuusinen et al. (2016). The albedo values measured for coniferous tree species in this study were similar to those measured by Lukeš et al., 2013a; Roberts et al., 2004; Hollinger et al., 2010; Kuusinen, 2014; Roberts et al., 2004). Albedo in deciduous plantations was clearly higher than that of the evergreen coniferous plantations, which is in agreement with Anderson (2011) and Breuer et al. (2003). The lower albedo in conifer trees might be explained by darker leaf color and needle-shaped leaves.
Our study confirmed that ground-based measurements of albedo allow estimating in-situ forest surface albedo with an albedo-meter setup similar as used by Riihelä and Manninen (2008).
Our study is in line with previous research on albedo changes (e.g., Betts, 2000; Bala et al., 2007; Pongratz et al., 2009) and advocates for considering albedo and radiative forcing consequences of changes in land use (Schwaiger and Bird, 2010) and for urban design (Akbari and Menon, 2009). Depending on colour, brightness and the type of land cover change, changes in albedo can have negative or positive consequences on the climate (Schwaiger and Neil Bird, 2010). This topic should be more widely considered to investigate the warming or cooling effects of trees on their environment.
4.2 Carbon and nitrogen stocks
Our data showed that tree species can have substantial effects on soil carbon and nitrogen stocks. Total C contents were higher in the broad-leaved stands, in particular alder and oak, while total N content in the alder stand was highest above all other stands, at 22 years plantation age. These findings somewhat disagree with Kooch et al. (2019). The changes in the C of the top soils in the different forest types might be caused by differences in quantity and quality of the forest floor inputs (Zheng et al., 2008). In fact, forest floor quality was the most important source of soil C in the forest ecosystem and may have an ambiguous relationship to tree canopy cover (Poeplau and Don, 2013).
Decomposition of the forest floor under coniferous trees is very slow. This may be related to poorer substrate quality of conifer needles compared to broadleaved litter (Paul et al., 2002). In this research, cypress had rather similar soil C and N stocks as broad-leaved trees, with the exception of alder. Thus, our study suggests that some broad-leaved plantations (i.e. alder) may have higher soil C stocks as coniferous plantations (cypress).
Significant differences in the soil carbon and nitrogen stocks between various tree stands support earlier findings highlighting the importance of tree species (Templer et al., 2012; Demessie et al., 2012; Sariyildiz et al., 2015; Berthrong et al., 2009; Li et al., 2012) and the effect of land cover on soil C and N (Franzluebbers and Stuedemann, 2002; Sreekanth et al., 2013).
The difference in C or N stocks in the mineral soil layers is largely regulated by species difference between forest floor fall inputs and outputs (decomposition), which is mainly controlled by forest floor quality and quantity (Laik et al., 2009; Li et al., 2012; Poeplau and Don., 2013) and biochemical properties between coniferous and deciduous species (Sariyildiz et al., 2015, Sariyildiz & Anderson 2003).
Soil N stocks followed the same trend of C and significant differences (p < 0.01) were detected between alder and other plantations. This is probably related to decomposition of forest floor (Kooch et al., 2019), In this study, decomposition of forest floor from alder plantation is apparently slow, which leads to more C and N accumulation than in other stands.
Our observations that mineral soil in broad-leaf stands (e.g., alder and oak stands) contained more C and N than in the conifer stand (e.g., cypress stand) disagrees with recent studies reporting higher C stocks in coniferous stands (Augusto et al. 2002; Jandl et al. 2014, Jandl et al. 2007). Other studies, however, provide evidence that soil C stocks in broad-leaved stands may be also higher than in conifers (Sariyildiz et al. 2015).
Our results further confirm other studies demonstrating a strong relationship between C and N stocks in mineral soil (De Vries, 2009; Sariyildiz et al., 2015), since the alder stand with the highest amount of C stocks also had the highest amount of N stocks. The high N stock in the alder stand, which is likely related to nitrogen fixation by alder roots, agrees with the findings of Sariyildiz and Anderson (2005), but contradicts Assad et al. (2013).
Soil C and N concentrations had a close relationship with soil organic matter (SOM) (Kooch et al., 2019). High SOM may be caused by continuous input of organic materials to the soil through forest litterfall (Gama-Rodrigues et al., 2010).
4.3 Regression analysis
The best selection principle of auxiliary variable is that the correlation between the auxiliary variable and the main variable is high, and the auxiliary variable is easily obtained (Wang et al., 2013). Among the soil variables, C stocks showed high correlation with soil N stocks (R2 0.73). In turn, the observed strong correlation between soil C and N suggest that C stocks can be used as a proxy for soil N stocks. Interestingly, soil C and N contents decreased with increasing soil clay content, which agrees with McLauchlan (2006).
Regression analysis showed a significant and very tight correlation between albedo and reflection. Climatic factors such as albedo depend on surface reflection and strongly was influenced by the type of the land cover. In our study sites, albedo was strongly influenced by various tree species of broad-leaved and coniferous. Our study showed that forestry activities such as plantation can not only substantially change soil C and N stock, but also the canopy albedo.
4.4 TOPSIS ranking
This study reports on the results of field measurements of albedo and soil carbon and nitrogen stocks in five different tree species in Hyrcanian forests at the North of Iran. Climate change management cannot be improved in an efficient manner, unless management strategies consider the sum of all effective factors on climate such as carbon stocks, albedo and other greenhouse gases emissions (Hovi et al., 2017). The present study shows how C and N stocks, canopy albedo can be combined to detect the optimum tree species with the best impact on mitigating global warming. The results of soil carbon and nitrogen stocks were combined with albedo into a decision-making algorithm to make inferences on which tree species would maximize the overall result, specifically the ones related to mitigate climate change consequences.
Planting trees as a climate mitigation measure has been challenged in various areas using coniferous or broad-leaved tree species ad planting material. Thus, improving our understanding of the factors controlling albedo, C and N stocks are fundamental for forest management activities, such as selection of tree species in plantation systems and design of reforestation systems for sustainable forests development under global warming conditions.
According to the TOPSIS technique ranking considering albedo and carbon and nitrogen stocks, alder was identified as the first priority of tree species to be used for forest development in northern Iran, followed by oak, maple, poplar and cypress. Thus, our study suggests that broad-leaved trees in this area should be favoured for forest development compared to coniferous tree species like cypress. This result is important for forest management decisions about reforestation and plantation for mitigating climate changes and global warming impacts. Future research should integrate climate change effects into assessment of the optimal use of forested lands for climate change mitigation.
Significant differences in albedo among tree species strongly support the hypothesis that broad-leaved tree species have higher albedo than coniferous trees. The soil C of alder and oak was higher than conifer tree stand, while soil N content only in the alder stand was higher than conifer tree stand. This may be related to the N-fixing property of alder, which has led to the higher N stocks than other stands.
5. Conclusions
Our findings demonstrated that surface albedo and soil carbon and nitrogen stocks under similar climatic and physiographic conditions are substantially changed by tree species in North of Iran. This observation may be used to evaluate the role of vegetation on global warming. Forest development with various tree species can help to reduce negative consequences of climate change. Until now, forest management has been mostly driven by economic consideration and timber production. We hope that in the future management decisions like proposed by this study will help mitigate climate change. For instance using conifer trees such as cypress may decrease the albedo, while broad-leaved trees such as alder appear to increase the albedo as well as soil C and N stocks. Reforestation with suitable native broad-leaved trees such as alder should help rehabilitate degraded natural forests and mitigate the climate change consequences in the Northern Iran.
References
Akhani H., Djamali M., Ghorbanalizade A., Ramezani E. (2010) Plant biodiversity of Hyrcanian relict forests, in Iran: an overview of the flora, vegetation, palaeoecology and conservation. Pak. J. Bot., Special Issue (S.I. Ali Festschrift) 42: 231-258.
Akbari H and Menon S. (2009) Global cooling: increasing world-wide urban albedos to offset CO2. Volume 94, Issue 3–4: 275–286.
Akin T., Özdemir N. (2003) Soil bulk density as related to soil particle size distribution and organic matter content Original scientific paper Izvorni znanstveni članak.
Akselsson C., Berg B., Meentemeyer V., Westling O. (2005). Carbon sequestration rates in organic layers of boreal and temperate forest soils—Sweden as a case study. Global Ecol. Biogeogr. 14, 77–84.
Allison L.E. (1975) Organic carbon in: ‘Methods of soil analysis’(ed) American Society of Agronomy, Part 2: Madison, Black, pp 1367–1378.
Anderson R.G. (2011), Biophysical considerations in forestry for climate protection Frontiers ecol. Environ.9174–82.
Anonymous (2015) Forest management planning in Pahnekola district of Sari. Organization of Forest and Rangelands and Watershed Management, Tehran, p 285.
Anonymous (2011) Forest management planning in Mahdasht district of Sari. Organization of Forest and Rangelands and Watershed Management, Tehran, p 301.
Aponte C., García-Fernández LV., Marañón T. (2013) Tree species effects on nutrient cycling and soil biota: A feedback mechanism favouring species coexistence. Forest Ecology and Management 2013(309):36. DOI:10.1016/j.foreco.2013.05.035.
Assad E., Pinto H.S., Pavão E., Luna R., Camargo P.B., Martinelli L.A., Martins S.C., Groppo J.D., Salgado P.R., Evangelista B., Vasconcellos E., Sano E.E. (2013) Changes in soil carbon stocks in Brazil due to land use. Paired site comparisons and a regional pasture soil survey, Biogeosciences, 10, 6141-61600, doi: 10.5194/bg-10-6141-2013.
Angelsen A and Wertz-Kanounnikoff S. (2008) What are the key design issues for REDD and the criteria for assessing options? Issues, options and implications, 11-22. CIFOR, Bogor, Indonesia.
Atlaskina K., Berninger F., Leeuw G de. (2015) Satellite observations of changes in snow-covered land surface albedo during spring in the Northern Hemisphere. The Cryosphere Discuss., 9, 2745–2782, 2015.doi:10.5194/tcd-9-2745-2015.
Alkama R and Cescatti A. (2016) Biophysical climate impacts of recent changes in global forest cover, Science, 351(6273), 600-604. doi:10.1126/science.aac8083.
Augusto L., Ranger J., Binkley D., Rothe A.(2002) Impact of several common tree species of European temperate forests on soil fertility. Annals of Forest Science 59(3), 233-253.DOI: https://doi.org/10.1051/forest:2002.020.
Bala G., Caldeira K., Wickett M., Phillips T. J., Lobell B.D., Delire C., Mirin A. (2007), Combined climate and carbon-cycle effects of large-scale deforestation, Proc. Natl. Acad. Sci. U.S.A.,104(16), 6550–6555.
Betts A.K and Ball J.H. (1997) Albedo over the boreal forest. Journal of Geophysical Research 102: 28901–28909.http://dx.doi.org/10.1029/96JD03876.
Berthrong S.T., Piñeiro G., Jobbágy E., Jackson R.B. (2012) Soil C and N changes with afforestation of grasslands across gradients of precipitation and plantation age. Ecological Applications 22(1):76-86 DOI: 10.2307/41416745.
Blake G.R. and Hartge K.H. (1986) Bulk density. In: Klute, A., Ed., Methods of Soil Analysis, Part 1—Physical and Mineralogical Methods, 2nd Edition, Agronomy Monograph 9, American Society of Agronomy—Soil Science Society of America, Madison, 363-382.
Bremner J.M., Mulvaney C.S. (1982). Nitrogen - total. In: Page, A.L., Miller, R.H., Keeney, R.R. (Eds.), Methods of Soil Analysis. second ed. American Society of Agronomy, Part 2, Madison, W.I., pp. 595–624.
Bright R.M, Antón-Fernóndes C., Astrup R., Cherubini F., Kvalevåg M., Strømman A.H., Peters G.P. (2014) Climate change implications of shifting forest management strategy in a boreal forest ecosystem of Norway. Global Change Biology 20: 607-621. http://dx.doi.org/10.1111/gcb.12451.
Bonan G.B. (2008) Forests and climate change: forcing feedbacks, and the climate benefits of forests. Science 320, 1444–1449. doi:http://dx.doi.org/10.1126/science.1155121.
Bouyoucos G.J. (1962) Hydrometer method improved for making particle size analysis of soils. J Agrobiol 56:464–465.
Breuer L., Eckhardt K., Frede H.G. (2003) Plant parameter values for models in temperate climates, Ecol. Model., 169, 237–293.
Burakowski E.A., Ollinger S.V., Lepine L., Schaaf C.B., Wang Z., Dibb J.E, Hollinger D.Y, Kim J., Erb A., Martin M. (2015) Spatial scaling of reflectance and surface albedo over a mixed-use, temperate forest landscape during snow-covered periods. Remote Sensing of Environment (158): 465–477.
Canadell J.G and Raupach M.R. (2008) Managing forests for climate change mitigation. Science 320(5882):1456–1457.
Davies-Barnard T., Valdes P. J., Singarayer J. S., Wiltshire A and Jones C. D. (2015) Quantifying the relative importance of land cover change from climate and landuse in the representative concentration pathways Glob. Biogeochem. Cycles 298 : 42–53.
De Vries F.T. (2009) Soil fungi and nitrogen cycling. Causes and consequences of changing fungal biomass in grasslands. Ph.D thesis Wageningen Universiteit, Wageningen 2009. – With ref. – With summary in English and Dutch. ISBN 978-90-8585-325-1.
De Vries W., Reinds G.J., Gundersen P., Sterba H. (2006) The impact of nitrogen deposition on carbon sequestration in European forests and forest soils. Global Change Biol. 12, 1151–1173.
Demessie A., Singh B.R., Lal O. (2012) Effects of eucalyptus and coniferous plantations on soil properties in Gambo District, southern Ethiopia.Acta Agriculturae. Scandinavica, Section B — Soil and Plant Science 62(5): 455-466. DOI: http://dx.doi.org/10.1080/09064710.2011.644575.
Dib A.E., Johnson C.E., Driscoll C.T., Fahey T.J., Hayhoe K. (2014) Simulating effects of changing climate and CO2 emissions on soil carbon pools at the Hubbard Brook Experimental Forest. Global Change Biology 20(5): 1643–1656. DOI: https://doi.org/10.1111/gcb.12436.
Ellison D., Cindy E., Bruno M., Bruno L., Caroline A., Sullivan. (2017) Trees, forests and water: Cool insights for a hot world. Global Environmental Change 43(51):51-61. DOI10.1016/j.gloenvcha.2017.01.002.
Finzi A.C., van Breemen N., Canham C.D. (1998) Canopy tree-soil interactions within temperate forests: Species effects on soil carbon and nitrogen. Ecol. Appl. 8, 440–446.
Franzluebbers A.j and Stuedemann J.A. (2002) Particulate and non-particulate fractions of soil organic carbon under pastures in the Southern Piedmont USA. Environmental Pollution Volume 116, Supplement 1, March 2002, Pages S53-S62.
Gama-Rodrigues E.F., Nair P.K.R., Nair V.D., Gama-Rodrigues A.C., Baligar V.C., Machado R.C.R. (2010) Carbon storage in soil size fractions under two cacao agroforestry systems in Bahia, Brazil. EnvironManag 45:274–283. doi:10.1007/s00267-009-9420-7.
Goodale C.L., Apps M.J., Birdsey R.A., Field C.B., Heath L.S., Houghton R.A., Jenkins J.C., Kohlmaier G.H., Kurz W., Liu S., Nabuurs G.J., Nilsson S., Shvidenko A.Z. (2002) Forest carbon sinks in the northern hemisphere. Ecological Applications 12(3): 891–899. doi: 10.1890/1051-0761(2002)012 [0891:FCSITN]2.0.CO;2
Hollinger D.Y., Ollinger S.V., Richardson A.D., Meyers T.P., Dails D.B., Martin M.E., Scott N.A., Arkebauer T.J., Baldocchi D.D., Clark K.L., Curtis P.S., Davis K.J., Desai A.R., Dragoni D., Goulden M.L., Gu L., Katul G.G., Pallardy S.G., Paw K.T., Schmid H.P., Stoy P.C., Suyker A.E., Verma S.B. (2010) Albedo estimates for land surface models and support for a new paradigm based on foliage nitrogen concentration. Global Change Biology 16, 696–710.
Hovi A., Liang J., Korhonen L., Kobayashi H., Rautiainen M. (2016) Quantifying the missing link between forest albedo and productivity in the boreal zone. Biogeosciences, 13, 6015–6030.
Hwang C.L and Yoon K. (1981) Multiple Attribute Decision Making: Methods and Applications. Springer-Verlag, New York, 260 p.
IPCC: Climate change (2007). The physical science basis, Cambridge University Press: Cambridge/New York, NY, 2007.
Jana B.K., Biswas S., Sonkar S., Majumder M., Roy P.K., Mazumdar A. (2009) Soil Carbon and Soil Respiration Rate of Urban Forests in Lower Gangetic Plain. International Journal of Applied Agricultural Research ISSN 0973-2683 Volume 4 Number 2 (2009) pp. 103–113.
Jandl R., Rodeghiero M., Martinez C., Cotrufo M.F., Bampa F., Wesemael B., Harrison R.B., Guerrini I.A., Richter D., Rustad L., Lorenz K., Chabbi A., Miglietta F. (2014) Current status, uncertainty and future needs in soil organic carbon monitoring. Science of The Total Environment 468–469: 376–383. https://doi.org/10.1016/j.scitotenv.2013.08.026.
Kindermann G., Michael O., Brent S., Jayant S., Kenneth A., Ewald R., Bernhard S., Sven W., Robert B. (2008) Global cost estimates of reducing carbon emissions through avoided deforestation. Proceedings of the National Academy of Sciences 105, no. 30 (2008): 10302-10307.
Kooch Y., Rostayee F., Hosseini S.M. (2014) Soil quality indices in pure and mixed forest stands of southern Caspian region. Ecopersia 3(2): 987-1001.
Kooch Y., Sanji R., Tabari M. (2019) The effect of vegetation change in C and N contents in litter and soil organic fractions of a Northern Iran temperate forest. Catena 178: 32–39.
Kaul M. (2010) Carbon budgets and carbon sequestration potential of Indian forests. PhD thesis, Wageningen University, Wageningen, The Netherlands. 178p. Available at [access date: 19.07.2017]: https://www.wur.nl/en/show/Carbon-budgets-and-carbon-sequestration-potential-of-Indian-forests.htm.
Kuusinen N., Lukeš P., Stenberg P., Levula J., Nikinmaa E., Berninger F. (2014) Measured and modelled albedos in Finnish boreal forest stands of different species, structure and understory. Ecological Modelling 284: 10–18.
Kuusinen N., Stenberg P., Korhonen L., Rautiainen M., Tomppo E. (2016) Structural factors driving boreal forest albedo in Finland. Remote Sensing of Environment 175 (2016) 43–51.
Larionova A.A., Rozanova L.N., Evdokimov I.V., Ermolaev A.M. (2002) Carbon budget in natural and anthropogenic forest-steppe ecosystems, Pochvovedenie 2, pp 177–185
Lal R., Kimble J.M., Follet R.F., Stewart B.A. (2001) Assessment methods for soil carbon advance in soil science. 1 st edn. CRC Press, Boca Raton, FL. (9781566704618).
Lal R. (2005) Forest soils and carbon sequestration. Forest Ecology and Management 220.242-258.
Li J.H., Jiao S.M., Gao R.Q., Bargett R.D. (2012) Differential effects of legume species on the recovery of soil microbial communities, and carbon and nitrogen contents in abandoned fields of the Loess Plateau, China. Environ Manage 50: 1193–1203.
Lovett, G.M.,Weathers, K.C., Arthur, M.A. (2002). Control of nitrogen loss from forested watersheds by soil carbon:nitrogen ratio and tree species composition. Ecosystems 5, 712–718.
Lovett G.M., Weathers K.C., Arthur M.A., Schultz J.C. (2004) Nitrogen cycling in a northern hardwood forest: do species matter? Biogeochemistry 67, 289–308.
Lukeš P., Stenberg P., Rautiainen M., Móttus M., Vanhatalo K. (2013a) Optical properties of leaves and needles for boreal tree species in Europe. Remote Sens. Lett. 4: 667–676. doi:10.1080/2150704X.2013.782112.
Lukeš P., Stenberg P., Rautiainen M. (2013b) Relationship between forest density and albedo in the boreal zone. Ecol. Modell. 261–262, 74–79.
Lara-Romero J., Campos-García J., Dasgupta-Schubert N., Borjas-García S., Tiwari DK., Paraguay-Delgado F., Jiménez-Sandoval S., Alonso-Nuñez G., Gómez-Romero M., Lindig-Cisneros R., Reyes De la Cruz H., Villegas J.A. (2017), Biological effects of carbon nanotubes generated in forest wildfire ecosystems rich inresinous trees on native plants. PeerJ 5:e3658; DOI 10.7717/peerj.3658.
Lutz D.A and Howarth R.B. (2014) Valuing albedo as an ecosystem service: implications for forest management. Clim. Chang. 124 (1–2), 53–63. http://dx.doi.org/10.1007/ s10584-014-1109-0.
Marcos E., Calvo L., Marcos J.A., Taboada A., Tárrega R. (2010) Tree effects on the chemical topsoil features of oak, beech and pine forests. Eur J For Res 129:25–30.
McLauchlan K.K. (2006) Effects of soil texture on soil carbon and nitrogen dynamics after cessation of agriculture. Geoderma 136, 289–299.
Mote T.L. (2008) On the role of snow cover in depressing air temperature. Journal of Applied Meteorology and Climatology, 47, 2008–2022.
Myhre G., Shindell D., Bréon F.M., Collins W., Fuglestvedt J., Huang J., Koch D., Lamarque J.F., Lee D., Mendoza B., Nakajima T., Robock A., Stephens G., Takemura T and Zhang H. (2013) Anthropogenic and Natural Radiative Forcing. In Stocker, T.F., D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex and P.M. Midgley, Eds., Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York: Cambridge University Press.
Nik H and Nik N. (2015) Climate Change And The Risk Messages Of Environment: A Visual Social Semiotics Analysis Of The Malaysian And German Online Newspapers Publication date 2015/12/2.
Naudts K., Chen Y., McGrath M.J., Ryder J.,Valade A., Otto J., Luyssaert S. (2016) Europe’s forest management did not mitigate climate warming. Science: Vol. 351, Issue 6273, pp. 597-600.DOI: 10.1126/science.aad7270.
Olson D.L. (2004) Comparison of Weights in TOPSIS Models. Mathematical and Computer Modelling.
Osher L.J., Matson P.A., Amundson R. (2003). Effect of land use change on soil carbon in Hawaii. Biogeochemistry 65: 213-232. - doi: 10.1023/A: 1026048612540.
Parsapour M.K., Kooch Y., Hosseini S.M., Alavi S.J. (2018). C and N cycle monitoring under Quercus castaneifolia plantation. Forest Ecology and Management 427 (2018) 26–36.
Prescott C.E and Vesterdal L. (2013) Tree species effects on soils in temperate and boreal forests: emerging themes and research needs. In: Forest Ecology and Management. Vol. 309. pp. 1-3.
Paul K.I., Polglase P.J., Nyakuengama J.G., Khanna P.K. (2002) Change in soil carbon following afforestation. Forest Ecol Manag 168: 241-257.
Poeplau C and Don A. (2013) Sensitivity of soil organic carbon stocks and fractions to different land-use changes across Europe. Geoderma Volume 192, January 2013, Pages 189-201.
Pongratz J., Raddatz T., Reick C. H., Esch M and Claussen M. (2009), Radiative forcing from anthropogenic land cover change since AD 800, Geophys. Res. Lett.,36, L02709, doi:10.1029/2008GL036394.
Roberts D.A., Ustin S.L., Ogunjemiyo S., Greenberg J., Dobrowski S.Z., Chen J., Hinckley T.M (2004) Spectral and structural measures of northwest forest vegetation at leaf to landscape scales. Ecosystems 7, 545–562.
Riihelä A. and Manninen T. (2008) Measuring the vertical albedo profile of a subarctic boreal forest canopy. Silva Fennica 42(5): 807–815.
Rautiainen M. and Stenberg P. (2005) Application of photon recollision probability in simulating coniferous canopy reflectance. Remote Sensing of Environment, 96, 98–107.
Shyur H. (2006). COTS Evaluation Using Modified TOPSIS and ANP. Applied Mathematics and Computation, 177 (1), pp. 251-259.
Smith P., Davies C.A., Ogle S., Zanchi G., Bellarby J., Bird N., Boddey R.M., MCNamara N.P., Powlson D., Cowie A., Noordwijk M., Davis S.C, Richter, D.D.B., Kryzanowski, L., van Wijk, M.T., Stuart, J., Kirton, A., Eggar, D., Newton-Cross G., Adhya T.K., Braimoh A.K. (2012) Towards an integrated global framework to assess the impacts of land use and management change on soil carbon: Current capability and future vision. Global Change Biology 18(7): 2089–2101. DOI: https://doi.org/10.1111/j.1365-2486.2012.02689.x.
Smolander S. and Stenberg P. (2005) Simple parameterizations for the radiation budget of uniform broad-leaved and coniferous canopies. Remote Sensing of Environment, 94, 355–363.
Sohngen B. and Mendelsohn R. (2003) An Optimal Control Model of Forest Carbon Sequestration. American Journal of Agricultural Economics. 85(2): 448-457.
Stephens G.L, O’Brien P.J., Webster P., Pilewski S., Kato., Li j. (2015) The albedo of Earth, Rev. Geophys., 53, 141–163, doi:10.1002/2014RG000449.
Streck C. (2012) Financing RE DD+: matching needs and ends. Current Opinion in Environmental Sustainability, 4(6), pp. 628–637. Available at: http://linkinghub.elsevier.com/retrieve/pii/S1877343512001376.
Sreekanth N.P., Shanthi PrabhaV., Padmakumar B., Thomas A.P. (2013) Effect of landuse conversion on soil carbon storage in a tropical grassland. Annals of Environmental Science / 2013, Vol 7, 101-112.
Scheidel A., Work C. (2016) Large-scale forest plantations for climate change mitigation? New frontiers of deforestation and land grabbing in Cambodia. International Institute of Social Studies (ISS)Kortenaerkade. An international colloquium 4-5 February 2016 Colloquium Paper No. 11.
Sariyildiz T., Anderson J.M. (2005) Variation in the chemical composition of green leaves and leaf litters from three deciduous tree species growing on different soil types. Forest Ecology and Management 210(1-3): 303-319. DOI: https://doi.org/10.1016/j.foreco.2005.02.043.
Sariyildiz T., Savaci G., Kravkaz I.S. (2015). Effects of tree species, stand age and landuse
change on soil carbon and nitrogen stock rates in northwestern Turkey. iForest 9: 165-170. – doi: 10.3832/ifor1567-008 [online 2015-06-18]
Schwaiger H.P and Bird D.N. (2010). Integration of albedo effects caused by land use change into the climate balance: Should we still account in greenhouse gas units? Forest Ecology and Management 260: 278–286.
Templer P.H., Mack M.C., Chapin F.S., Christenson L.M., Compton J.E., Crook H.D., Currie W.S., Curtis C.J., Dail D.B., D'Antonio C.M., Emmett B.A., Epstein H.E., Goodale C.L., Gundersen P., Hobbie S.E., Holland K., Hooper D.U., Hungate B.A., Lamontagne S., Nadelhoffer K.J., Osenberg C.W., Perakis S.S., Schleppi P., Schimel J., Schmidt I.K., Sommerkorn M., Spoelstra J., Tietema A., Wessel W.W., Zak D.R. (2012) Sinks for nitrogen inputs in terrestrial ecosystems: a meta-analysis of 15N tracer field studies. Ecology 93 (8): 1816-1829. DOI: https://doi.org/10.1890/11-1146.1.
Thiel S., Ammannato L., Bais A., Bandy B., Blumthaler M., Bohn B., Engelsen O, Gobbi GP, Gröbner J., Jakel E., Junkermann W., Kazadzis S., Kift R., Kjeldstad B., Kouremeti N., Kylling A., Mayer B., Monks P.S., Reeves C.E., Schallhart B., Scheirer R., Schmidt S., Schmitt R., Schreder J., Silbernagl R., Topaloglou C., Thorseth T.M., Webb A.R., Wendisch M., Werle P. (2008) Influence of clouds on the spectral actinic flux density in the lower troposphere (INSPECTRO): overview of the field campaigns, Atmos. Chem. Phys., 8, 1789– 1812, doi:10.5194/acp-8-1789-2008.
Uzun S. and Yildirim B.F. (2016) Equipment selection in ship building process: TOPSIS, MOORA, VIKOR application. Eurasian Academy of Sciences Eurasian Business & Economics Journal. Volume: S2 .113-124.
Vesterdal L., Schmidt I.K., Callesen I., Nilsson L.O., Gundersen P. (2008). Carbon and nitrogen in forest floor and mineral soil under six common European tree species. Forest Ecology and Management 255: 35-48. - doi: 10.1016/j.foreco. 2007.08.015.
Wang K., Zhang C.R., Li W.D. (2013). Predictive mapping of soil total nitrogen at a regional scale: A comparison between geographically weighted regression and Cokriging. Applied Geography, 42(8): 73–85. doi: 10.1016/j.apgeog.2013.04.002.
Yatso K.N and Lilleskov E.A. (2016) Effects of tree leaf litter, deer fecal pellets, and soil properties on growth of an introduced earthworm (Lumbricus terrestris): Implications for invasion dynamics. Soil biology and biochemistry 94.p :181-190. https://doi.org/10.1016/j.soilbio.2015.11.030.
Zheng H., Ouyang Z., Xu W., Wang X., Miao H., Li X., Tian Y. (2008) Variation of carbon storage by different reforestation types in the hilly red soil region of southern China. Forest Ecol Manag 255: 1113-1121.



