Schlüsselbegriffe: Fagus orientalis, lineare Regression, Standortindex, Boden, physiographische Faktoren
Abstract
This study examined the relationships between soil, climatic and physiographic variables and site productivity in Fagus orientalis Lipsky stands in Artvin, Northeastern Turkey. The site productivity described by the site index (SI, top height at stand age of 100 years) was available for three forest locations. SI varied significantly depending on the edaphic, climatic and physiographic variables. A linear regression model was constructed to test the hypothesis that soil, climatic and physiographic variables could be used to predict the SI of the Fagus orientalis forest types in the study region. The most important single variable in the model was precipitation in the driest quarter of the year, which accounted for 55.1% of the variability in SI and was positively correlated. In a multiple regression model, precipitation in the driest quarter, radiation index, slope and physiological soil depth were the most important variables for explaining SI in the study region. 80% of the observed SI variation was explained by these variables. The SI models developed in this study provide a basis for understanding the complex relationship between forest productivity and site conditions. The findings of this study will assist resource managers in making cost-effective decisions about the management of Fagus orientalis forests in the northeastern region of Turkey.
Zusammenfassung
Diese Studie untersuchte die Beziehungen zwischen klimatischen, physiographischen und edaphischen Variablen und Standortproduktivität in Fagus-orientalis-Lipsky-Beständen in Artvin im Nordosten der Türkei. Die Standortproduktivität wurde durch den Site Index (SI, Oberhöhe bei Bestandesalter von 100 Jahren) beschrieben und war an drei Waldorten verfügbar. Der SI variierte signifikant in Abhängigkeit von den klimatischen, physiografischen und edaphischen Variablen. Ein lineares Regressionsmodell wurde zur Testung der Hypothese entwickelt, dass klimatische, physiografische und edaphische Variablen verwendet werden können, um SI bei Fagus-orientalis-Beständen in der Untersuchungsregion vorherzusagen. Die wichtigste einzelne Variable im Modell war der Niederschlag im trockensten Viertel des Jahres, der 55,1 % der SI-Variabilität erklärte und positiv korreliert war. In einem multiplen Regressionsmodell waren der Niederschlag im trockensten Viertel, der Strahlungsindex, die Hangneigung und die physiologische Bodentiefe die wichtigsten Variablen zur Erklärung der Standortproduktivität. 80 % der beobachteten SI-Variation wurden durch diese Variablen erklärt. Die in dieser Studie entwickelten SI-Modelle bieten eine Grundlage für ein Verständnis der komplexen Beziehung zwischen Waldproduktivität und Standortbedingungen im Untersuchungsgebiet. Die Ergebnisse dieser Studie werden Ressourcenmanager dabei unterstützen, kosteneffektive Entscheidungen für die Bewirtschaftung von Fagus-orientalis-Wäldern in der nordöstlichen Region der Türkei zu treffen.
1 Introduction
The determination of site productivity is fundamental for effective forest resource management. Estimating productivity at the early stages of stand development allows for defining appropriate silvicultural treatments to be applied in subsequent years. Site productivity depends on environmental factors such as physiographic, edaphic, and climatic conditions and is often represented in forestry by site index (SI) or stand dominant height at a predetermined reference age. Forest site productivity is usually estimated using site and stand factors (Bueis et al., 2016). SI is generally used as an indicator of the forest productivity, since SI is strongly correlated with wood production (Ortega and Montero, 1988; Skovsgaard and Vanclay, 2008). One crucial issue when estimating forest site productivity is the relationship between environmental factors and tree height growth (Szwaluk and Strong, 2003).
Forests are specific areas, including many interconnected environmental factors that maintain tree growth and affect plant development (Afif-Khouri et al., 2011). Tree growth is the outcome of the effects of genetic characteristics and the environment. The environmental factors affecting the growth and development of trees are physiography (slope, aspect, altitude, and slope position), soil (physical and chemical soil properties) and climate characteristics (precipitation, temperature, moisture). Changes in these environmental conditions can affect the development of forest trees and stands. The changes in forest growth may be species-specific and depend on site fertility and environmental conditions at the stand level (Kellomäki et al., 2008; Ge et al., 2013; Granda et al., 2013; Torssonen et al., Rohner et al., 2016; Seiler et al., 2017; Koulelis et al., 2022). The productivity of stands composed of the same tree species in the same geographical region can be quite different, if local characteristics are variable. The productivity of the forest site is known as the natural capacity of a forest area affected by the existing growth factors (light, temperature, water, nutrients) and is usually expressed as the tree height at a certain age (Skovsgaard and Vanclay, 2008). Thus SI is often regarded as a key measure of the productivity of commercial forests (Socha, 2008; Palahí et al., 2008; Bontemps and Bouriaud, 2014; Yılmaz et al., 2015; Subedi and Fox, 2016).
Climate determines the distribution of tree species and shapes forest composition. On large scales precipitation has been identified to affect species richness (Ter Steege et al., 2006), composition (Hall and Swaine, 1976) and species occurence (Engelbrecht et al., 2007; Swaine 1996; Toledo et al., 2012). On smaller scales, the physiographic and edaphic characteristics have also been reported to affect the distribution of tree species (John et al., 2007). The growth of tree species in the sites depends on water, nutrients and light as primary sources (Toledo et al., 2012). The changes in environmental factors may cause the same tree species to grow differently in the same region but under different site conditions. Topography may influence climate in a particular region. Climate and soil are presumably among the most important factors determining growth and species distributions (Walthert and Meier, 2017).
This study was carried out in three locations within a distinct geographic region, where climate elements (precipitation, temperature, moisture) were variables depending on the topography (Kantarcı, 1995). The effect of climatic, edaphic and physiographic variables on site productivity was investigated for natural Fagus orientalis stands in the Artvin region of Northeastern Turkey. Fagus orientalis is one of the most important broadleaf tree species due to its productivity, high growing stock and wide distribution in Turkey (Anonymous, 2015). Fagus orientalis, one of twelve beech species, naturally occurs in the Balkan countries, including Bulgaria, Greece, Turkey, the Caucasus region and northern Iran (Kandemir and Kaya, 2009).
The relationships between SI and site characteristics may provide useful information for forest management.
2 Material and methods
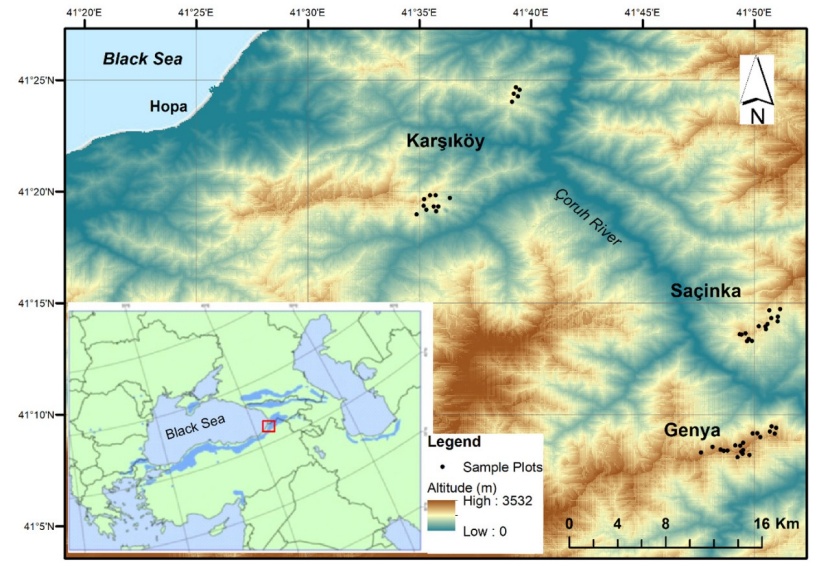
Figure 1: Distribution range of oriental beech (Kandemir and Kaya 2009) and location of studied sites. / Abbildung 1: Verbreitungsgebiet der Orientbuche (Kandemir and Kaya 2009) und Lage der untersuchten Standorte.
2.1 Study area
The study area is located in the Artvin Region in Northeastern Turkey (Fig. 1). The sample plots were taken from pure Fagus orientalis stands in Karşıköy-Erenköy region (Karşıköy, from now on) and in the Genya and Saçinka regions. Artvin region is in a transitional climate between the land influenced by moist air from the Black Sea and the Inner Black Sea land. Genya mountain and Saçinka hill receive additional moisture from the Black Sea. While some of these moist air masses reach the research area along the Çoruh valley from Batum, some of them reach the research area by passing over the Cankurtaran (900 m) passage between Sultan Selim Mountain (1500 m) and Balıklı mountain (1890 m). Air masses that cross the Cankurtaran pass rise leave a certain amount of precipitation on the slopes facing the sea. The research area in the Karşıköy region is fed by both the air masses passing over Cankurtaran pass and the air masses coming from the Çoruh valley. Therefore, this area is very humid. The air masses moving south from Borcka are moving in the direction of Murgul and some along the Çoruh valley. The air masses that continue along the Çoruh valley rise on the slopes of the Genya and Saçinka hill cool down and release the moisture in them. Compared to Karşıköy, Genya and Saçinka ridges receive less rainfall (Table 1). With the effect of this moist air, Fagus orientalis can still establish pure stands on the slopes of Genya and Saçinka, allthough these stands are on the margins of this species.
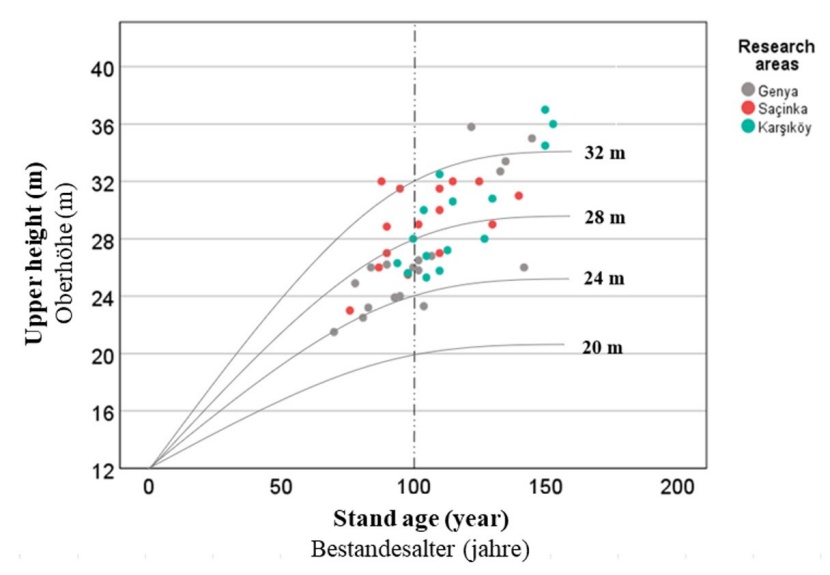
Figure 2: Site index curves of the yield table used (Carus 1998) and the age and height values of the sample plots. / Abbildung 2: Bestandesalter und Oberhöhen der Probenflächen und Site-Index-Kurvem aus Ertragstafeln (Carus 1998).
2.2 Sample Plots
Twenty, fifteen and fifteen sample plots were established in Genya, Saçinka and Karşıköy, respectively. The sample plots were rectangular in shape, and their sizes varied between 400 m2 under > 70% crown closure and 600 m2 when crown closure ranged between 41–70%. The trees in the sample plots were numbered following a clockwise direction and the diameter of breast height (DBH) 1.3 m above ground was measured with a caliper. Tree height was measured on the 100 highest trees per hectare using a Blume-Leiss device and the stand age was quantified by counting the annual rings froman increment core taken at height of 0.3 m above ground. The stand top height was then defined as the average total height of the 100 highest trees per hectare.
Stand upper height and stand age values of the sample plot are shown on the site index curves of the used yield tables (Fig. 2).
The site index (SI) was determined by using the yield table developed by Carus (1998) for even-aged beech forests for a reference age of 100 years. The altitude (m), aspect (degree) and slope (%) of the sampling points were measured with an altimeter, compass and inclinometer, respectively. The aspect variable was converted into a topographic solar-radiation index (TRASP) used by Roberts and Cooper (1989) for statistical evaluation. Trasp is calculated by the following equation:
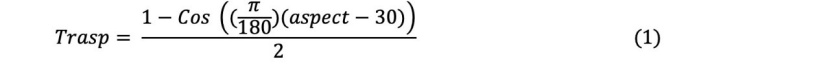
This conversion assigns value “0” to the field in the north-northeast direction (the coldest and most humid direction) and value “1” to the warmer, drier slopes in the south-southwest direction (Moisen and Frescino, 2002).
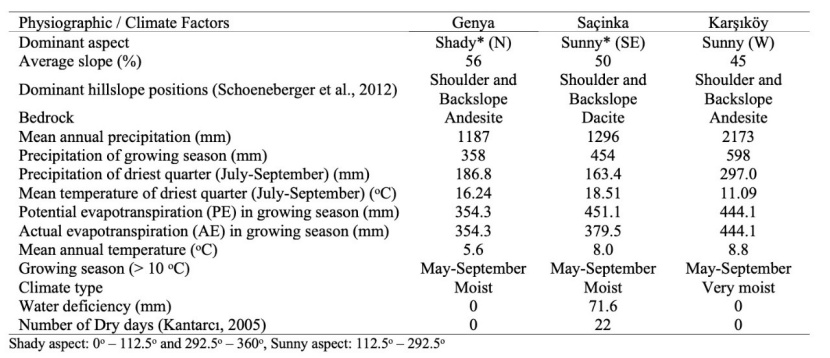
Table 1: Selected climatic and physiographic features of the research areas. / Tabelle 1: Ausgewählte klimatische und physiografische Merkmale des Untersuchungsgebiets.
2.3 Soil Characteristics and Climate
In the sample plots, soil pits were dug (down to maximum 1.20 m depending on the depth of the bedrock), and absolute (ASD) and physiological soil depths (PSD) were determined (Kantarcı, 2000). In the soil profile, mineral soil horizons were separated and samples were collected (Kantarcı, 1995). The fine earth weight (gr l-1, < 2 mm size) (FP) and the amount of skeleton (gr l-1, ≥ 2 mm size) (SKE) of soil horizons were determined according to Corti et al. (1998). Fine earth refers to soil particles smaller than 2 mm in diameter. Skeleton defines soil rock fragments or soil coarse fraction, that is, particles with 2 mm as a minor diameter (Miller and Guthrie, 1984).
The particle size analysis of soil samples was performed according to the Bouyoucos hydrometer method and the soil types were determined according to international texture classes (Gülçur, 1974). Organic soil carbon was determined by using the Walkley-Black wet burning method. Soil organic matter was estimated by multiplying soil organic carbon by 1.72 (Kantarcı, 2005). Soil pH was measured on a 1/2.5 soil: deionized water suspension according to Page et al. (1982). The available water capacity (AWC) was calculated based on the differences in field capacity moisture (FC) and wilting point moisture (WP) values determined in soil moisture device with pressure table (Karaöz, 1989). The available water capacity calculated in percentage for the horizons was correlated with fine earth weight and calculated in mm (Kantarcı, 2005).
Climate data (monthly average precipitation, monthly average temperature, and monthly maximum temperature) obtained from Paleoclim were used for some bioclimatic variables and Thornthwaite climate analysis. The bioclimatic variables data under current conditions (average 1979–2013) with a spatial resolution of 30 arc second (~1 km) was obtained from Paleoclim (https://chelsa-climate.org/downloads/; accessed on 2 October 2021) (Karger et al., 2017). These variables are often used in species distribution models and ecological studies. The bioclimatic variables used in the study were annual mean temperature (AT), max temperature of the warmest month (TWM), mean temperature of the driest quarter (TDQ), mean temperature of the coldest quarter (TCQ), annual precipitation (AP), precipitation of the wettest month (PWM), precipitation of the driest month (PDM), precipitation of the driest quarter (PDQ) and precipitation of the warmest quarter (PWQ).
The selected areas were within the borders of the Artvin province and characterized by topographical differences affecting the climate conditions. Climate analysis was performed for the research regions (Thornthwaite, 1948) (Table 1). In Thornthwaite climate analysis, potential evapotranspiration (PE) is calculated by the following formulas.
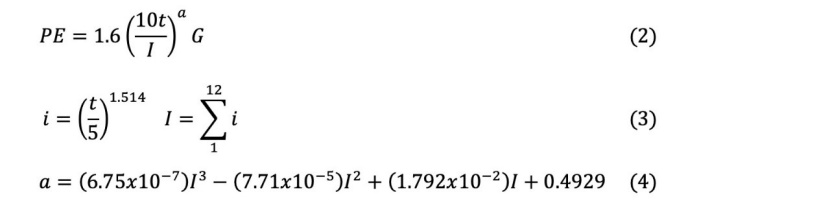
2.4 Statistical Analyses
SI as the dependent variable, and physiographic factors (slope, aspect, altitude, slope position), climatic characteristics, and soil characteristics changes as independent variables were tested by the analysis of variance. Pearson correlation and multiple regression analysis were implemented among them, aiming to predict SI for all research regions. SPSS statistics version 26 was used for the analyses (SPSS Inc., Chicago IL, USA).
3 Results
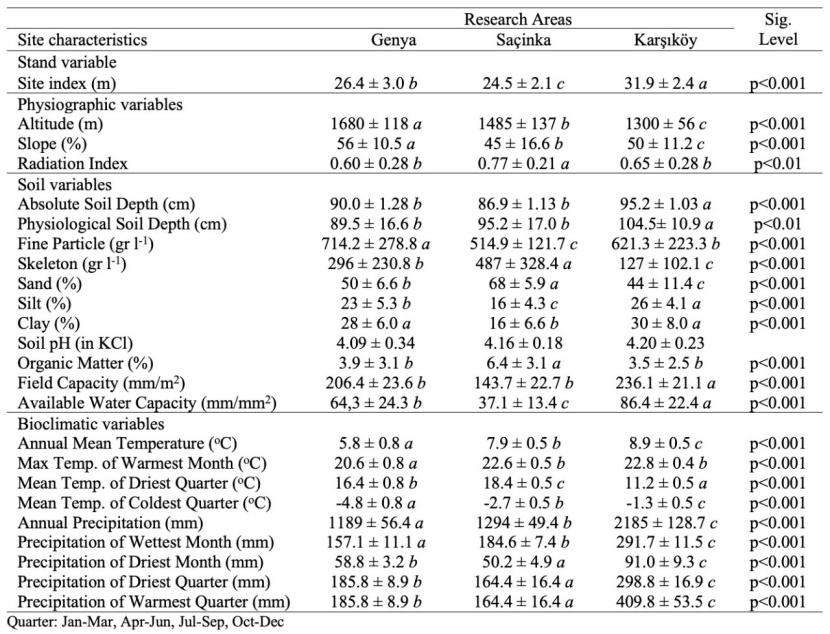
Table 2: Variance analysis results of the entire research area. / Tabelle 2: Ergebnisse der Varianzanalyse des gesamtes Untersuchungsgebietes.
The variance analysis suggest that the research areas were different according to the main soil properties (sand, silt and clay, fine earth, skeleton, field capacity, available water capacity). While physiological soil depth was similar in the Genya and Saçinka regions, the amounts of organic matter were similar in the Karşıköy and Genya regions. Physiographic factors and climatic characteristics also differed according to the regions (Table 2).
The Pearson correlation analysis indicated significant relationships between site index and soil, climatic and physiographic variables (Fig. 3).
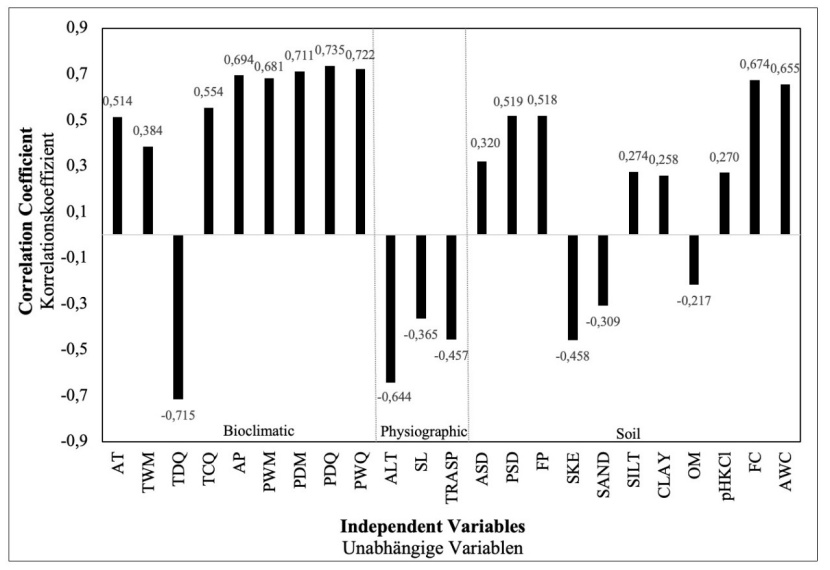
Figure 3: Correlations between site index and environmental variables*. / Abbildung 3: Korrelationen zwischen Site Index und Umweltvariablen.
(* Environmental variables are the sum of bioclimatic, physiographic and soil variables, AT:Annual mean temperature, TWM:Max. temperature of warmest month, TDQ:Mean temperature of driest quarter, TCQ:Mean temperature of coldest quarter, AP:Annual precipitation, PWM: Precipitation of wettest month, PDM:Precipitation of driest month, PDQ:Precipitation of driest quarter, PWQ:Precipitation of warmest quarter, ALT:Altitude, SL: Slope, TRASP: Topographic solar-radiation index, ASD: Absolute soil depth , PSD: Physiological soil depth, FP: Fine particle, SKE: Skeleton, OM: Organic matter, FC: Field capacity, AWC: Available water capacity)
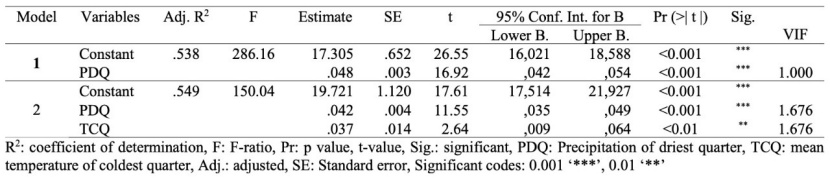
Table 3: Prediction models for SI using only climate variables. / Tabelle 3: Vorhersagemodelle für SI nur mit Klimavariablen.
For the bioclimatic variables, SI showed a negative correlation with TDQ (r=−0.715, p<0.001) and a positive correlation with AT (r=0.514, p<0.001), TWM (r=0.384, p<0.001), TCQ (r=0.554, p<0.001), AP (r=0.694, p<0.001), PWM (r=0.681, p<0.001), PDM (r=0.711, p<0.001), PDQ (r=0.735, p<0.001), PWQ (r=0.722, p<0.001). For the physiographic variables, SI had a negative correlation with ALT (r=-0,644, p<0.001), SL (r=−0.365, p<0.001), TRASP (r=−0.457, p<0.001) (Fig. 3). For the soil variables, SI had a positive correlation with ASD (r=0.320, p<0.001), PSD (r=0.519, p<0.001), FP (r=0.518, p<0.001), SILT (r=0.274, p<0.001), CLAY (r=0.258, p<0.001), pH(KCl) (r=0.270, p<0.001), FC (r=0.674, p<0.001) and AWC (r=0.655, p<0.001), a negative correlation SKE (r=-0.458, p<0.001), SAND (r=-0.309, p<0.001) and OM (r=−0.217, p<0.001) (Fig. 3).
Multiple regression analysis was performed using the variables of climatic variables, soil properties and physiographic factors with a high correlation with site productivity values (Table 3, Table 4). Multicollinearity was determined among some bioclimatic variables. The variables with VIF values >10 were removed from the regression model (Senaviratna and Cooray, 2019).
At regression analysis, according to bioclimatic variables, two regression models were obtained for the site index. The prediction percentages of regression models ranged from 53.8 to 54.9 (Table 3). The results of the regression model (Step 2) showed that the probability site index could be predicted by climatic variables such as PDQ and TCQ (Table 3).
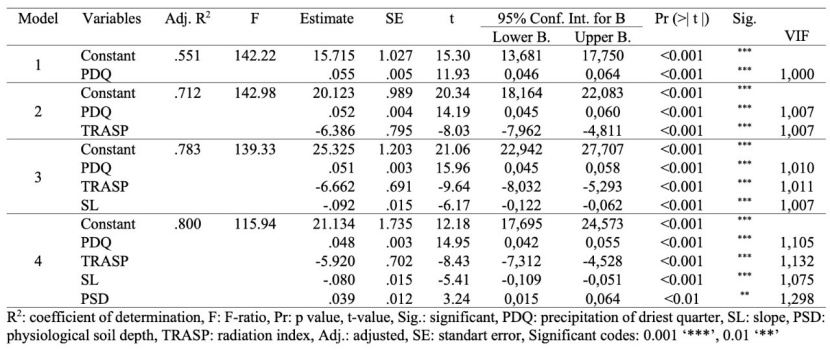
© Table 4: Prediction of SI according to environmental variables. / Tabelle 4: Vorhersagemodelle für SI mit allen getesteten Umweltvariablen.
At regression analysis, according to all variables, four regression models were predicted for the site index. The prediction percentages of regression models ranged from 55.1 to 80.0 (Table 4). The results of the regression model (Step 4) indicated that the probability site index could be predicted by such soil, climatic and physiographic variables as PDQ, TRASP, SL and PSD (Table 4).
4 Discussion
4.1 Effects of physiographic factors on site productivity
Altitude is one of the most important ecological factors that affect the productivity of forest trees and the species composition of the forest, along with the other physiographic factors (slope, aspect, slope position), that change the climatic characteristics (precipitation, temperature, solar radiation). With this effect, different site conditions may occur in the same geographical region. As it increases, the vegetation period is shortened, and site productivity decreases. The literature states that the site index decreases depending on the increase in altitude (Klinka et al., 1996; Corona et al., 1998; Socha, 2008; Pinno et al., 2009; Sharma, 2012; Yılmaz et al., 2015). In the study, the highest site index value (31.9 m) was measured in the Karşıköy region, where the average altitude is the lowest. In this study, concerning the selected study areas, we found a negative correlation between the altitude and the site index (p<0.001, r=-0,644) (Fig. 3). In this respect, our results are compatible with previous studies. The lowest site index value was measured in the Oriental beech forests in the Saçinka region, where the average altitude is 1475 m. It may seem contradictory that the site index was higher in the Genya region, where the altitude was higher, compared to the Saçinka region. However, this can be explained depending on the ecology of the Oriental beech and other ecological factors. Oriental beech prefers north-facing slopes that are deep, well-drained, moist and sloping and rich in water and nutrients, and it may reach up to 2000 m altitude.
The sample plots in the Saçinka region were taken from the soils developed from dacite bedrock (Table 1). While the sand content (68%) and the amount of skeleton (487 gr l-1) of the soils developed from dacite bedrock were high, the amount of fine particles, clay (16%) and silt (16%) were low (Table 2). These soil properties led to adverse conditions in terms of water availability and available water capacity. The average available water capacity was the lowest. In the Saçinka region, the land slope degree was lower compared to the Genya region, and the dominant aspect was the sunny aspect. The average temperature and average annual temperature during the growing period were higher (Table 1), which increased the evapotranspiration rate in the region, causing a water deficit during the growing period. Even though the soil depths of the research areas are close to each other and deep (>85 cm), site productivity in the Saçinka region was the lowest due to low water-holding capacity and available water capacity. The sample plots in the Genya region were more productive than the sample plots in the Saçinka region since they have sufficient soil depth and soil moisture in shady aspects. During our field studies, we occasionally observed intensive fog formation on Oriental beech forests on Genya ridges, although it did not turn into precipitation. It is also known that Oriental beech prefers the sites where soil depth is sufficient, drainage is good, and relative humidity of the air is high (Yılmaz, 2019). Therefore, Karşıköy, where the site index was highest, is more suitable than the two research regions in terms of the variables described above. Research areas were similar concerning the vegetation period (Table 1).
The sample areas in the Genya region are in the shady aspect (north-facing slopes), and the dominant aspects of the sample areas in the Saçinka and Karşıköy regions are sunny (south-facing slopes). Since the aspect factor will change the physical and chemical soil properties of the site along with other physiographic factors, it is expected to affect the productivity index of the Oriental beech. As explained above on altitude, the combined effect of the aspect with slope is significant on-site characteristics. The combined effect of slope and aspect is highly effective in the growth of trees by causing changes in solar radiation (Waring, 2000). In the correlation analysis, there was a positive correlation between site index and transformed aspect variable (TRASP) (Fig. 3). Steinbrenner (1965) and Corona et al. (2008) concluded a positive relationship between aspect and site index. The aspect plays a key role in determining the temperature regime of any site (Bennie et al., 2008). In the Northern Hemisphere, south-facing slopes receive much more solar radiation than other directions and are relatively hot and dry, which also leads to differences in the growth of forests, depending on the ecological requirements of the tree species (Rydgren et al., 2003; Saremi et al., 2014). Oriental beech prefers high air humidity and moist soils (Odabaşı et al., 2004) and is less developed on south-facing slopes, where soil moisture exposed to more solar radiation will be low, compared to north-facing slopes. Considering that Oriental beech is a species with high demand for humidity and temperature, precipitation in the driest month or the driest quarter is expected to influence the site index. The region with the lowest precipitation in the driest month and the driest quarter is the Saçinka region, and the region with the highest precipitation is the Karşıköy region (Table 3). In the driest month and the driest quarter, the site index value is the lowest in the Saçinka region, where the precipitation is the lowest. In the correlation analysis, there was a strong positive relationship between the site index value and the precipitation in the driest month and the precipitation in the driest quarter (Fig. 3, p<0.001, r=0,711, r=0.735). The relationship between the maximum temperature of the warmest month and the site index was positive. Similar results were reported by Monserud et al. (2006).
Along with other physiographic factors, the slope of the ground modifies the amount of solar radiation received by the surface and has been identified as an important factor in determining the ecological conditions at a site. Solar radiation is the dominant component of the surface energy balance and influences ecologically critical factors of microclimate, including near surface temperatures, evaporative demand and soil moisture content (Bennie et al., 2008). The fact that these critical factors are far from optimum may negatively affect the nutrition of plants and the activity of soil organisms and may also disrupt physical and chemical decomposition. Stony, low-moisture and shallow soils develop in high-slope sites with similar topographic and pedological characteristics. Therefore, the productivity of forest trees is low in these areas. In this study, there was a negative correlation between slope degree and site index (Fig. 3). Our results are confirmed by the literature (Klinka et al., 1996; Corona et al., 1998; Günlü et al., 2006; Anwar, 2007; Socha, 2008; Sharma, 2012).
The research areas were similar in terms of slope (hillslope) position. All the sample areas were taken from backslope and shoulder. Soils with different characteristics developed due to different slope, aspect, altitude, bedrock and climatic characteristics. The site productivity of the Oriental beech forests growing on these soils was different (Table 2). The soils of the Karşıköy and Genya region developed from andesite bedrock are loamy clay type, while the soils of the Saçinka region developed from dacite bedrock of sandy clay loam-sandy loam type. The main reason for the development of different types of soil in the Saçinka region is the difference in bedrock. Soils with higher amounts of sand develop from the dacite bedrock, which contains quartz in its composition. In general, the soils dominated by sand are poor in terms of nutrient and water holding capacity. With the effects of both bedrock characteristics and climate (temperature, precipitation etc.) and topography (slope, aspect) affecting physical and chemical decomposition, the soil skeleton per unit volume was higher, and the fine earth weight per unit volume was lower in the soils of the Saçinka region (Table 2). Water and nutrient storage capacities of the soils with high skeleton and low amount of fine earth weight are low. The site productivity was, therefore, low in the Saçinka region, where the soils with these characteristics are located.
4.2 Effects of climate variables on site productivity
Site productivity depends on environmental conditions (biotic, edaphic, and climatic) associated with a particular location (Clutter et al., 1983) and is directly influenced by changes in temperature and precipitation regimes (Latta et al., 2010). Our results indicated that climate variables (temperature and precipitation) are important in determining site productivity for Oriental beech. Site index was related to all selected climate variables. The SI values ranged between 22.30 and 34.46 m, indicating a large variation in productivity among the different sites. As expected, the highest productivity was observed in the Oriental beech stands located in the area with the highest precipitation and highest annual average temperature. The increase in precipitation amounts of the warmest month and the warmest quarter and annual precipitation increased site productivity. While the correlation between average temperature, annual precipitation and site productivity is positive, the temperature of the driest month is negatively related to site productivity. Considering the climate variables associated with SI, parameters related to temperature and precipitation affect the presence of water in the soil. An increase in site productivity can be expected with an increase in average temperature in humid and very humid climatic regions. However, increasing the temperature in the dry and warm months increases evaporation, and site productivity may decrease. When evaporation increases, water availability for trees decreases, and there is a decrease in site productivity (Paulo et al., 2014). The positive correlation between precipitation amounts (annual precipitation and precipitation amount in the driest months) and site productivity is consistent with the findings of previous studies. Similarly, the site index of forest trees was reported to have a positive relationship with annual precipitation and precipitation of the driest month (Güner et al., 2016; Özel et al., 2020). Besides the studies in which the site productivity increased with the increase in the annual precipitation (Watt et al., 2010; Sabatia and Burkhart 2014), there are many studies in which precipitation is used in the modeling of the site index (Ung et al., 2001; Chen et al., 2002; Seynave et al., 2005; Seynave et al., 2008; Albert and Schmidt, 2010).
The productivity indexes of Oriental beech were different by region. Since the climatic and physiographic characteristics of the regions change, the soil characteristics of the site change, and thus site productivity also differs. Soil, which has an important role in the growth of forest trees, determines the distribution of tree species along with climatic characteristics. The Karşıköy region receives the highest annual average precipitation. Climatic factors, such as precipitation and temperature, place primary controls on the growth and development of natural vegetation (Rundquist and Haarington, 2000). An increase can be expected in site productivity, along with an increase in the amount of precipitation. In general, tree growth increases with precipitation (Dauber et al., 2005; Mondek, 2021). However, precipitation during the vegetation period is more decisive. The highest precipitation in the vegetation period occurs in the Karşıköy region. Carter and Klinka (1990) and Corona et al. (1998) also determined a higher site index in the regions where precipitation was higher in annual and growing periods. Our results show that site productivity increases with the increase in annual total precipitation and in growing season precipitation.
4.3 Effects of soil properties on site productivity
Some of the soil properties result from environmental conditions, depending on the characteristics of the bedrock where the soils are formed. The presence and availability of water in the soil affect the site productivity depending on climate variables (Watt et al., 2010). The available water capacity of the soils is 13.9%, 9% and 7.2%, respectively, in Karşıköy, Genya and Saçinka. While the average site index was highest in Karşıköy region, with the highest available water capacity and field capacity, it was the lowest in the Saçinka region. In the correlation analysis performed for the whole research area, significant positive correlations were found between the available water capacity and field capacity values of the soils and site productivity (Fig. 3). These results are consistent with other studies (Carter and Klinka, 1990; Yılmaz et al., 2015; Eckhart et al., 2019).
The fact that available water capacity and field capacity are highly correlated with site productivity means that the presence of water is a key element for growth (Falkenmark and Rockström, 2006). The water stress that will occur due to lack of water in the soil leads to stomatal closure and a decrease in carbon retention efficiency. In conclusion, a reduction in cambial activity and foliage growth occurs, followed by a decrease in wood production. The site index in the Saçinka region, where there was water deficit during the growing period, was lower. Stape et al. (2004) observed that an increase in water availability increases quantum efficiency, leaf area and decreases carbon allocation to roots. Given the importance of water availability for plant growth, a range of variables related to soil moisture, available water for stands, or precipitation has been tested before on several models (Snowdon et al., 1999; Pinjuv et al., 2006; Ritchie and Hamann, 2008).
Soil depth is important because the deeper the soil, the more space available for roots. As the soil depth increases, the area available to the tree roots expands, and the water and nutrient storage capacity of the soil increases. Physiological soil depth (PSD), the actual soil depth available to tree roots, is positively correlated with site productivity (Schmidt and Carmean, 1988; Turvey et al., 1990). Limited soil volume on shallow sites can limit site productivity because shallow soils have lower soil volume and, therefore, lower total water holding capacity. In the research areas, physiological soil depths varied between 92-104 cm, and deep and very deep soils are dominant. While the Saçinka and Genya regions are similar in terms of physiological soil depth, the Karşıköy region is different and the highest. In the correlation analysis performed for all research areas, a positive correlation was found between the physiological soil depth and site productivity (Fig. 3). In the research area, where there was sufficient soil depth for the development of trees, the soil depth difference of 8–12 cm showed its effect on-site productivity. Considering the whole area where tree roots can spread, this depth difference is a considerable value. The increase in soil depth is important in terms of increasing the volume of soil in which tree roots can develop and hold and the water and nutrients to be kept in the soil. For these reasons, it is a variable used in determining site productivity (Pyatt et al., 2001). The increase in absolute and physiological depths of soils increases the site productivity, and physiological soil depth was included in the site index prediction model (Table 3). The positive correlation between site productivity and soil depths is mentioned in a large number of studies (Yılmaz, 2005; Mckenney and Pedlar, 2003; Günlü et al., 2006).
PDQ, TRASP, SL, and PSD variables explained 80.0% of the variation in the SI of Oriental beech. Although it does not participate in the regression model, the ALT factor has a significant effect on the site index since it affects climate variables and soil properties. It changes soil moisture conditions and growth by affecting climatic characteristics, such as temperature and precipitation. It plays an indirect role in litter decomposition and nutrient supply. It shows all these effects together with aspect, another physiographic factor because aspect controls the soil moisture conditions, the activity of soil organisms, soil temperature and photosynthesis with the effect of solar radiation. TRASP is included in the model. It could be said that Oriental beech develops better in deeper and stone-free soils where AWC is higher. If the site is similar to other conditions, the increase in precipitation positively affects the presence of water in the soil. Climate variables, such as the amount of PDQ and TDQ, are also very effective on the site productivity of Oriental beech. The variation in stand height growth of Oriental beech trees in this study could be explained using the climate variables PDQ and TDQ. These variables are very intuitive and can be easily interpreted when explaining the growth of any tree species. The inclusion of climatic variables together with soil properties and physiographic characteristics significantly improved the fit statistics of the stand height growth model. These results are consistent with the findings of previous studies. For example, Wang et al. (2007) and Bravo-Oviedo et al. (2008) reported that including climate variables improved the fit and predictive accuracy of the model in their studies. The topographic (Anwar, 2007; Oviedo et al., 2011; Farrelly et al., 2011; Mohamed et al., 2014) and edaphic (Fonte et al., 2003; Ercanli et al., 2008) variables were generally included in site index prediction models.
5 Conclusion
The results of this study provide information on the productivity of Oriental beech stands in Northeast Turkey. In this study, significant correlations between the site productivity of Oriental beech forests and topographic factors (ALT, SL, TRASP), soil properties (SKE, AWC, FC, PSD e.g.) and climate variables (AT, AP, TWM, TDQ, TCQ, PDQ, PWQ, PWM, PDM) were determined. In the regression analysis, it was concluded that important variables affecting the productivity of Oriental beech stands in Northeast Turkey were one soil variable (PSD), three physiographic properties (SL and TRASP), and two climate variables (PDQ and TDQ). These variables together explained the change in site index by 80%. Considering the variables included in the model, it could be said that low precipitation, low temperature, and poor nutrients limit the growth in Oriental beech forests, respectively. The models developed in this study can be used to estimate stand height at reference age, given height at a point in time with relevant climate variables, edaphic variables, and physiographic factors. In addition, since height estimated at the reference age will provide the site index value, the models can also serve as site index equations. Oriental beech forests have different site productivity in the regions that get the sea effect and moderately get the sea effect in the Northeast of Turkey. In the regions moderately under the sea effect, the stand productivity is controlled by physiographic factors, climate variables and soil properties. In this region, the productivity of stands on soils developed from dacite bedrock in sunny aspects is lower than the productivity of stands on soils developed from basalt bedrock in shady aspects. The water deficit in the soil and, thus, the available water capacity is among the most important variables affecting productivity. In the Saçinka region, site productivity is the lowest due to water deficit (lowest AWC, FC, PDM, PWQ, and PDQ). Site productivity attracts considerable attention, especially when timber production is the primary goal, because it can present the production capacity of stands in advance. Depending on whether site productivity is low or high, management plans can be changed to optimize stand productivity. Stakeholders and forest administrations can use the results obtained from this study to plan the investments and work to achieve the best performance for forest production in sites with similar characteristics.
References
Afif-Khouri E., Alvarez-Alvarez P., Fernandez-Lopez M.J., Oliveria-Prendes J.A., Camara-Obregon A., 2011. Influence of climate, edaphic factors and tree nutrition on site index of chestnut coppice stands in north-west Spain. Forestry, Vol. 84, No. 4, doi:10.1093/forestry/cpr025
Albert M., Schmidt M., 2010. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst.) and common beech (Fagus sylvatica L.). For. Ecol. Manage. 259, 739–749.
Anonymous., 2015. Turkey Forest Asset. Republic of Turkey General Directorate of Forestry (in Turkish).
Anwar C., 2007. Prediction of teak Bonita (site index) in central java, Indonesia, based on soil properties. Indonesian Journal of Forestry Research, v. 4, n. 1, p. 9–18
Bennie J., Huntley B., Hıll A., Wıltshıre M.O., Baxter R., 2008. Slope, aspect and climate: spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecol Model 216, 47–59.
Bontemps J.D., Bourıaud O., 2014. Predictive approaches to forest site productivity: recent trends, challenges and future perspectives. Forestry, v. 87, p. 109–128.
Bravo-Oviedo A., Tomé M., Bravo F., Montero G., del Rio M., 2008. Dominant height growth equations including site attributes in the generalized algebraic difference approach. Can. J. For. Res. 38: 2348–2358.
Bueis T., Bravo F., Pando V., Turrión M.B., 2016. Relationship between environmental parameters and Pinus sylvestris L. site index in forest plantations in northern Spain acidic plateau. iForest 9: 394–401. – doi: 10.3832/ifor1600-008
Carter R.E., Klınka K., 1990. Relationships between growing-season soil water-deficit, mineralizable soil nitrogen and site index of coastal douglas fir. For Ecol Manag 30, 301–311.
Carus S., 1998. Increment and growth in even aged beech (Fagus orientalis Lippsky.) forests. PhD Thesis, Istanbul University, Istanbul, Turkey (in Turkish)
Chen H.Y.H., Krestov P.V., Klinka K., 2002. Trembling aspen site index in relation to environmental measures of site quality at two spatial scales. Can. J. For. Res. 32, 102–119.
Clutter J.L., Fortson J.C., Pienaar L.V., Brister G.H., Bailey R.L., 1983. Timber Management: A Quantitative Approach. Krieger Publishing Company, Malabar, FL.
Corona P., Scotti R., Tarchiani N., 1998. Relationship between environmental factors and site index in douglas fir plantations in Central Italy. For Ecol Manag, 110, 1–3, 195-207.
Corti, G., Ugolini, F. C., & Agnelli, A., 1998. Classing the soil skeleton (greater than two millimeters): proposed approach and procedure. Soil Science Society of America Journal, 62(6), 1620–1629.
Dauber E., Frederıcksen T.S., Peňa-Claros M., 2005. Sustainability of timber harvesting in bolivian tropical forests. For Ecol Manag 214, 294–304.
Eckhart T., Pötzelsberger E., Koeck R., Thom D., Laır G.L., Loo M., Hasenauer H., 2019. Forest stand productivity derived from site conditions: an assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Ann For Sci, 76, 1, 1–11. https://doi.org/10.1007/s13595-019-0805-3
Engelbrecht B.M., Comıta L.S., Condıt R., Kursar T.A., Tyree M.T., Turner B.L., Hubbell S.P., 2007. Drought sensitivity shapes species distribution patterns in tropical forests. Nature, 447, 80–82.
Falkenmark, M., and Rockström, J. 2006. The new blue and green water paradigm: Breaking new ground for water resources planning and management. Journal of water resources planning and management, 132(3), 129–132.
Farrelly N., Dhubhaın A.N., Nıeuwenhuıs M., 2011. Site index of Sitka spruce (Picea sitchensis) in relation to different measures of site quality in Ireland. Can J For Res 41, 265–278.
Ge Z.M., Kellomäki S., Peltola H., Zhou X., Väisänen H., 2013. Adaptive management to climate change for Norway spruce forests along a regional gradient in Finland. Climatic Change 118: 275–289. http://dx.doi.org/ 10.1007/s10584-012-0656-5
Gülçur F., 1974. Physical and chemical analysis methods of soil, Istanbul University, Faculty of Forestry, No. 201, 225 p. (in Turkish).
Güner Ş.T., Çömez A., Özkan K., Karataş R., Çelik N., 2016. Modelling the productivity of Anatolian black pine plantations in Turkey. J Fac For Istanbul Univ 66(1):159-172. https ://doi.org/10.17099 / jffiu .18731 (in Turkish).
Günlü A., Yılmaz M., Altun L., Ercanlı I., Küçük M., 2006. Correlationships Between Site İndex And Some Edaphic And Physiographic Factors Of Pure Oriental Spruce (Picea Orientalis Link.) in Artvin Genya Mountain, Suleyman Demirel University, Journal of Natural and Applied Sciences, 1, 1–10.
Granda E., Camarero J.J., Gimeno T.E., Martínez-Fernández J.,Valladares F., 2013. Intensity and timing of warming and drought differentially affect growth patterns of co-occurring Mediterranean tree species. European Journal of Forest Research 132(3): 469–480. http://dx.doi. org/10.1007/s10342-013-0687-0
Hall J.B., Swaıne M.D. 1981. Distribution and ecology of vascular plants in a tropical rain forest. Forest vegetation in Ghana. Geobotany 1, Junk Publishers, The Hague, 383 p.
John R., Dallıng J.W., Harms K.E., Yavıtt J.B., Stallard R.F., Mırabello M., Hubbell S.P., Valencıa R., Navarrete H., Vallejo M., Foster R.B. 2007. Soil nutrients influence spatial distributions of tropical tree species. PNAS 104 (3), 864–869. https://doi.org/10.1073/pnas.0604666104
Kandemir G., Kaya Z., 2009. EUFORGEN Technical Guidelines for genetic conservation and use of oriental beech (Fagus orientalis). Bioversity International, Rome, Italy, 6 p.
Kantarcı M.D., 1995. Regional Ecological Units in the Eastern Black Sea Region, I. National Black Sea Forestry Congress, 23–25 October, Proceedings Book, Volume: 3, 111–138, Trabzon.
Kantarcı M.D., 2000. Soil Science. Istanbul University, Faculty of Forestry, Second Edition, 420p (in Turkish).
Kantarcı M.D., 2005. Knowledge of Forest Ecosystems. İstanbul University Faculty of Forestry, İstanbul, 379 p. (in Turkish).
Karaöz M.Ö., 1989. Methods of Determination of Some Physical Properties of Soils related to Water Economy in the Laboratory, İstanbul University Journal of Forestry Faculty, 39, 2, 133–144 (in Turkish).
Karger, D.N., Conrad, O., Böhner, J., Kawohl, T., Kreft, H., Soria-Auza, R.W., Zimmermann, N.E., Linder, P., Kessler, M., 2017. Climatologies at high resolution for the Earth land surface areas. Scientific Data. 4 170122. https://doi.org/10.1038/sdata.2017.122
Kellomäki S., Strandman H., Nuutinen T., Peltola H., Korhonen K.T., Väisänen H., 2005. Adaptation of forest ecosystems, forest and forestry to climate change. FINADAPT. Working Paper 4. Finnish Environment Institute Mimeographs 334, Helsinki. https://helda.helsinki.i/ handle/10138/41042
Klinka K., Wang Q., Carter R.E., Chen H.Y.H., 1996. Height growth-elevation relationships in subalpine forests of interior British Columbia. Forestry Chronicle, 72, 193–198.
Koulelis P.P., Fassouli V.P., Petrakis P.V., Ioannidis K.D. Alexandris S. (2022). The impact of selected climatic factors on Greek fir growth on Mt Giona in mainland Greece based on tree ring analysis. Austrian Journal of Forest Science, 139. Jahrgang (2022), Heft 1, S. 1–30.
Latta G., Temesgen H., Adams D., Barrett T., 2010. Analysis of potential impacts of climate change on forests of the United States Pacific Northwest. For. Eco. Man. 259: 720–729.
Mckenney D.W., Pedlar J.H., 2003. Spatial models of site index based on climate and soil properties for two boreal species in Ontario, Canada. For Ecol Manag,175, 497–507. https://doi.org/10.1016/S0378-1127(02)00186-X
Miller, F. T., & Guthrie, R. L. 1984. Classification and distribution of soils containing rock fragments in the United States. Erosion and productivity of soils containing rock fragments, 13, 1–6.
Mohamed A., Reich R.M., Khosla R., Aguırre-Bravo C., Brıseño, M.M., 2014. Influence of climatic conditions, topography and soil attributes on the spatial distribution of site productivity index of the species rich forests of Jalisco, Mexico. Journal of Forestry Research, 25, 1, 87–95
Moisen G.G., Frescino T.S., 2002. Comparing five modelling techniques for predicting forest characteristics. Ecol Model 157, 2–3, 209–225. https://doi.org/10.1016/S0304-3800(02)00197-7
Mondek J., Matějka K., Gallo J., ProkůpkováA., Hájek V., 2021. Picea abies and Pseudotsuga menziesii radial growth in relation to climate: case study from South Bohemia. Austrian Journal of Forest Science, 138, 3, 209–244.
Monserud R.A., Huang S., Yang Y., 2006. Predicting lodgepole pine site index from climatic parameters in Alberta. For. Chron., 82, 562–571.
Odabaşı T., Çalışkan A., Bozkuş H.F., 2004. Forest Tending, İstanbul University Faculty of Forestry, İstanbul, 192 p. (in Turkish).
Ortega A., Montero G., 1988. Evaluación de la calidad de las estaciones forestales. Revisión bibliográfica. [Forest site quality assessment. A literature review]. Ecología 2: 155–184 (in Spanish).
Oviedo A.B., Montero G., 2005. Site index in correlation to edaphic variables in stone pine (Pinus pinea L.) stands in South West Spain. Ann For Sci 62, 1, 61–72 https://doi.org/10.1051/forest:2004086
Özel C., Güner Ş.T., Türkkan M., Akgül S., Şentürk Ö., 2020. Modelling the site index of Pinus pinaster plantations in Turkey using ecological variables. J. For. Res. 32 (2), 589-598 https://doi.org/10.1007/s11676-020-01113-x
Paulo J.M., Palma J.H.N., Gomes A.A., Faias S.P., Tome J., Tome M., 2014. Predicting site index from climate and soil variables for cork oak (Quercus suber L.) stands in Portugal. New For. 46, 293-307 https://doi.org/10.1007/s11056-014-9462-4
Pınno B.D., Paré D., Guındon L., Bélanger N., 2009. Predicting productivity of trembling aspen in the Boreal Shield ecozone of Quebec using differentsources of soil and siteinformation. For Ecol Manag 257, 782–789. https://doi.org/10.1016/j.foreco.2008.09.058
Pinjuv G., Mason E.G., Watt M., 2006. Quantitative validation and comparison of a range of forest growth model types. For Ecol Manag 236, 37-46. https://doi.org/10.1016/j.foreco.2006.06.025
Pyatt D.G., Ray D., Fletcher J., 2001. An ecological site classification for forestry in Great Britain. Forestry Commission Bulletin, 124, Forestry Commission, Edinburgh
Ritchie M.W., Hamann J.D., 2008. Individual-tree height-, diameter- and crown-width increment equations for young Douglas-fir plantations. New Forests 35,173–186.
Roberts D.W., Cooper S.V., 1989. Concepts and techniques of vegetation mapping. In Land Classifications Based on Vegetation: Applications for Resource Management. USDA Forest Service General Technical Report INT-257, Ogden, UT, p. 90–96
Rohner, B., Weber, P., Thürig, E. 2016. Bridging tree rings and forest inventories: How climate effects on spruce and beech growth aggregate over time. Forest Ecology and Management, 360:159–169.
Rundquist B.C., Harrington J.A., 2000. The effects of climatic factors on vegetation dynamics of tallgrass and shortgrass cover. Geocarto International, Vol. 15, No. 3. 33-38.
Rydgren K., Klandr H., Kland T., 2003. Species response curves along environmental gradients. A case study from SE Norwegian swamp forests. Journal of Vegetation Science 14, 6, 869-880. https://doi.org/10.1111/j.1654-1103.2003.tb02220.x
Senaviratna N.A.M.R., and Cooray T.M.J.A. 2019. Diagnosing multicollinearity of logistic regression model. Asian Journal of Probability and Statistics, 5(2), 1–9.
Saremi H., Kumar L., Turner R., 2014. Airborne LiDAR derived canopy height model reveals a significant difference in radiata pine (Pinus radiata D. Don) heights based on slope and aspect of sites. Trees, 28, 3, 733–744.
Schmidt M.G., Carmean W.H., 1988. Jack pine site quality in relation to soil and topography in north central Ontario. Can. J. For. Res., 18: 297–305.
Schoeneberger, P.J., D.A. Wysocki, and E.C. Benham, and Soil Survey Staff., 2012. Field book for describing and sampling soils. Version 3.0. U.S. Department of Agriculture, Natural Resource Conservation Service, Lincoln, NE.
(PDF) Digital Classification of Hillslope Position. Available from: https://www.researchgate.net/publication/271130831_Digital_Classification_of_Hillslope_Position [accessed Jun 18 2022].
Seiler, R., Kirchner, J.W., Krusic, P.J., Tognetti, R., Houliea, N., Andronico, D., Cullotta, S., Egli, M., D'Arrigo, R., Cherubini, P. 2017. Insensitivity of Tree-Ring Growth to Temperature and Precipitation Sharpens the Puzzle of Enhanced Pre-Eruption NDVI on Mt. Etna (Italy). PLoS ONE, 12(1): e0169297. https://doi: 10.1371/journal. pone. 0169297
Seynave I., Gégout J.C., Hervé J.C., Dhôte J.F., Drapier J., Bruno E. et al., 2005. Picea abies site index prediction by environmental factors and understorey vegetation: a two-scale approach based on survey databases. Can. J. For. Res. 35, 1669–1678.
Seynave I., Gégout J.C., Hervé J.C., Dhôte J.F., 2008. Is the spatial distribution of European beech (Fagus sylvatica L.) limited by its potential height growth? J. Biogeog. 35, 1851–1862.
Sharma R.P., Brunner A., Eid T., 2012. Site index prediction from site and climate variables for norway spruce and Scots pine in Norway. Scand J For Res 27, 7, 619–636. https://doi.org/10.1080/02827581.2012.685749
Skovsgaard J.P., Vanclay J.K., 2008. Forest Site Productivity: A Review Of The Evolution Of Dendrometric Concepts For Evenaged Stands. Forestry. 81, 13–31. https://doi.org/10.1093/forestry/cpm041
Snowdon P., Jovanovıc T., Booth T.H., 1999. Incorporation of indices of annual climatic variation into growth models for Pinus radiata. For Ecol Manag 117, 187–197. https://doi.org/10.1016/S0378-1127(98)00463-0
Socha J., 2008. Effect of topography and geology on the site index of picea abies in the West Carpathians, Poland. Scand J For Res 23, 203–213. https://doi.org/10.1080/02827580802037901
Stape J., Bınkley D., Ryan M.G., 2004. Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For Ecol Manag 193, 1–2, 17–31. https://doi.org/10.1016/j.foreco.2004.01.020
Steinbrenner E., 1965. The influence of individual soil and physiographic factors on the site index of Douglas-fir in Western Washington. In: Forest-soil relations (CT Youngberg, ed), Oregon State University Press, Corvallis, OR, USA. 261–270.
Subedi S., Fox T.R., 2016. Predicting loblolly pine site index from soil properties using partial least-squares regression. For Sci 62, 4, 449–456. https://doi.org/10.5849/forsci.15-127
Swaine M., 1996. Rainfall and soil fertility as factors limiting forest species distributions in Ghana. J Ecol, 84, 419–428. https://www.jstor.org/stable/2261203
Szwaluk K. S., Strong W.L., 2003. Near-surface soil characteristics and understory plants as predictors of Pinus contorta site index in southweatern Alberta, Canada. For Eco Man. 176, 13–24
Ter Steege H., Pıtman N.C.A., Phıllıps O.L., Chave J., Sabatıer D., Duque A., Molıno J.F., Prévost M.F., Spıchıger R., Castellanos H., Von Hıldebrand P., Vásquez P., 2006. Continental-scale patterns of canopy tree composition and function across Amazonia. Nature, 443, 444–447.
Thornthwaıte C.W., 1948. An approach toward a rational classification of climate. Geographical Review, 38, 1, 55–94.
Toledo M., Peňa-Claros M., Bongers F., Alarcón A., Balcázar J., Chuvıňa J., Leaňo C., Lıcona J.C., Poorter L., 2012. Distribution patterns of tropical woody species in response to climatic and edaphic gradients. J Ecol 100, 253–263. https://doi.org/10.1111/j.1365-2745.2011.01890.x
Torssonen P., Strandman H., Kellomäki S., Kilpeläinen A., Jylhä K., Asikainen A., Peltola H., 2015. Do we need to adapt the choice of main boreal tree species in forest regeneration under the projected climate change? Forestry 88: 564–572. http://dx.doi.org/10.1093/forestry/cpv023
Turvey N.D., Booth T.H., Ryan P.J., 1990. A soil technical classification for Pinus radiate (D. Don) plantations. II. A basis for estimation of crop yield. Aust. J. Soil Res., 28: 813–824.
Ung C.H., Bernier P.Y., Raulie, F., Fournier R.A., Lambert M.C., Re´gnie`re J., 2001. Biophysical site indices for shade tolerant and intolerant boreal species. For. Sci. 47, 83–95.
Walthert L., Meier E.S., 2017. Tree species distribution in temperate forests is more influenced by soil than by climate. Ecol Evol. 1–12.
Wang Y., LeMay V.M., Baker T.G., 2007. Modelling and prediction of dominant height and site index of Eucalyptus globulus plantations using a nonlinear mixed-effects model approach. Can. J. For. Res. 37: 1390–1403.
Waring R.H., 2000. A process model analysis of environmental limitations on the growth of Sitka spruce plantations in Great Britain. Forestry 73, 1, 65–79. https://doi.org/10.1093/forestry/73.1.65
Watt M.S., Palmer D.J., Kımberley M.O., Höck B.K., Payn T.W., Lowe D.J., 2010. Development of models to predict Pinus radiata productivity throughout New Zealand. Can J For Res 40, 3, 488–499. https://doi.org/10.1139/X09-207
Yılmaz M., 2005. Studies on some site factors affecting the growth (productivity) of the beech in pure oriental beech (Fagus orientalis Lipsky) ecosystems in East Karadeniz Region, 188 p., Phd thesis, Karadeniz Technical University, Trabzon (in Turkish).
Yılmaz M., 2019. Effects of soil and environmental factors on the site productivity of pure Oriental beech forests in Akkuş region of Turkey. Eurasian Journal of Forest Science, 7, 2, 107–120. https://doi.org/10.31195/ejejfs.527905
Yılmaz M., Usta A., Öztürk İ., 2015. Relationships between site ındices and ecological factors for black alder stands in Turkish eastern Black Sea region. Fresenius Environmental Bulletin, 24, 4, 1285–1293.



