Schlüsselbegriffe: Orientbuche, bioklimatische Zone, Generhaltung, phänotypische Variation
See below the issue 4/2022 as E-Paper or have a look at our E-Paper archive dating back to 1955.
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Abstract
Beech forests have a very important place not only in timber production, but also in environmental and soil protection, with a wide variety of provided benefits. This study has two main objectives: 1. to examine morphogenetic variations of seeds and seedlings of oriental beech (Fagus orientalis Lipsky) in relation to geoclimatic factors in the natural distribution of oriental beech in Turkey and 2. to provide basic information about gene conservation and management of oriental beech forests by morphogenetically examining the variations among and within 11 populations in the natural distribution of oriental beech in Turkey. There were statistical differences among and within the populations depending on seed and seedling characteristics measured in the study. A Penrose's analysis suggest that, the highest distance value was evident between Sinop-Abalı and Trabzon-Çaykara populations (Pn1,7: 3.490) and the lowest distance value between Trabzon-Maçka and Giresun- Kulakkaya populations (Pn6,8: 0.477). We conducted correlation analysis between bioclimatic factors and seed and seedling morphological characteristics and found positive correlations between mean of the minimum temperature of the coldest month and seedling length as well as the number of side branches of the one and two-year-old seedlings, while we found negative correlations between altitude and these characteristics. The seed sizes had statistically significant correlations with each other and with seedling length, root collar diameter and the number of side branches of the one and two-year-old seedlings. Since preservation of gene resources in-situ is important in terms of continuity of variation and conservation of biological diversity, preservation of existing populations and continuity of genetic variation should be ensured. In future forest establishment studies of oriental beech, we advise attention with regards to altitude zones and geographic distances. Using seedlings produced using seeds of a confirmed origin in such studies will likely increase both success and reduce losses in genetic diversity.
Zusammenfassung
Buchenwälder haben nicht nur in der Holzproduktion, sondern auch im Umwelt- und Bodenschutz einen sehr wichtigen Stellenwert und bieten vielfältige Vorteile. Diese Studie hat zwei Ziele: 1. Untersuchen der morphologischen Variationen von Samen und Sämlingen der Orientbuche (Fagus orientalis Lipsky) in Bezug auf geoklimatische Faktoren in der natürlichen Verbreitung der Orientbuche in der Türkei und 2. Sammeln von grundlegenden Informationen über die Generhaltung und das Management von Orientbuchenwäldern durch morphogenetische Untersuchung der Variationen zwischen und innerhalb von 11 Populationen in der natürlichen Verbreitung der Orientbuche in der Türkei. Es gab statistische Unterschiede zwischen und innerhalb der Populationen in Abhängigkeit von den in der Studie gemessenen Samen- und Sämlingsmerkmalen. Eine Penrose-Analyse zeigte den höchsten Distanzwert zwischen den Sinop-Abalı- und Trabzon-Çaykara-Populationen (Pn1,7: 3.490) und den niedrigsten Distanzwert zwischen den Trabzon-Maçka- und Giresun-Kulakkaya-Populationen (Pn6,8: 0,477). Die Ergebnisse einer Korrelationsanalyse zwischen bioklimatischen Faktoren und morphologischen Merkmalen von Samen und Sämlingen zeigten positive Korrelationen zwischen dem Mittelwert der Mindesttemperatur des kältesten Monats und den Werten der Höhe der Sämlinge sowie der Anzahl der Seitenzweige gefunden auf ein- und zweijährigen Sämlingen, aber negative Korrelationen zwischen der Seehöhe und diesen Merkmalen. Auch die Samengrößen zeigten statistisch signifikante Korrelationen untereinander sowie mit Sämlinglänge, Wurzelhalsdurchmesser und der Anzahl der Seitenäste der ein- und zweijährigen Sämlinge. Da die Erhaltung von Genressourcen in-situ im Hinblick auf die Kontinuität der Variation und die Erhaltung der biologischen Vielfalt wichtig ist, sollten die Erhaltung bestehender Populationen und die Kontinuität der genetischen Variation sichergestellt werden. Bei geplanten Aufforstungen der Orientbuche sind sowohl Höhenzonen als auch die Erhaltung des genetischen Variation zu beachten. Durch Verwendung von Setzlingen, die aus Saatgut mit bekannter Herkunft gezogen wurden, kann sowohl der Erfolg gesteigert als auch Verluste der genetischen Diversität reduziert werden.
1 Introduction
Forest establishment practices are expensive and long-term investments. In order to guarantee the future of these investments, it is necessary to use seeds and seedlings with superior genetic characteristics. Additionally, it is essential to know the population that is the source of the genetic-vegetative material from which the seedlings are produced and the genetic variations of the trees in the population in order to determine the genetic quality of a seedling. Conservation of genetic diversity is one of the basic components of sustainable forest management (Kaya 1990; Millar and Marshall 1991). Intraspecies genetic diversity is one of the most important factors in the resistance of forest trees to diseases and their adaptation to changing climatic conditions (Bilgen et al. 2011; Güney et al. 2014; Atar and Turna 2018; Güney et al. 2019; Atar et al. 2020).
Conservation of genetic diversity plays an important role in protecting species in the face of harsh ecological conditions. Also, the purpose of gene conservation is to develop conservation strategies and to maintain evolutionary potential (Alan 2017). Thus, populations with higher genetic diversity, to be an option against both adverse conditions that may occur and against future needs that may change, are a safeguard (St. Clair and Howe 2011). At the beginning of breeding programs, intraspecies genetic diversity studies are emphasized. The higher the genetic diversity of trees among and within the populations of a species, the greater the chances of geneticists to choose populations and genotypes suitable for their purposes. On the one hand, genetic diversity enables forest tree populations to overcome changes such as climate as well as diseases that occur in the environment, on the other hand, it can create an opportunity in tree breeding for many regions and usages with the evolution of the species (Ledig 1986; St. Clair and Howe 2011).
Local populations, which constitute a reserve against genetic contamination, should be protected to be used as genetic resources in the future (Işık 1988). In tree breeding, primarily natural stands are used as a genetic resource. According to natural selection theory, the gene pool is wide, so the natural range of species rich in genetic diversity is also wide (Yahyaoğlu and Genç 2007). It is stated that the best way to determine the genetic variation in a species is by comparing the populations in different habitats (Chmura and Rozkowski 2002; Güney et al. 2015; Atar et al. 2017). The species that spread over very large areas have a lot of geographic variation and local forms (Kaya 1990; Atar and Güney 2021; Atar 2021). Turkey's hilly geographical structure and changing climate over short distances and soil characteristics encourages the formation of local forms in forest tree populations even in short distances. Determining the genetic structure of natural forests, each of which is a gene resource, is important due to its economic, ecological and aesthetic features (Işık 1988; Güney et al. 2013). The spatial heterogeneity of the oriental beech (Fagus orientalis Lipsky) distribution in Turkey is an indicator of high genetic diversity within and between populations. In addition, in a study examining past, present and future distributions of oriental beech under climate change projections, pollen data indicate Turkey and the western Caucasus as refugia for oriental beech during the last glacial maximum (Dagtekin et al. 2020). Genetic diversity can be determined by a variety of techniques. This can be made with the help of morphological and biological characters or molecular markers. (Mettler and Gregg 1969; Suangtho et al. 1999; Yahyaoğlu and Genç 2007). Studies on the determination of genetic diversity in forest trees first started based on morphological characters (Ürgenç 1982; Doğan 1997). Heredity and environment have effects on morphological and physiological characteristics of forest trees. As an alternative to classical method in determining genetic diversity, studies are carried out at the molecular level including isoenzyme analysis, RAPD, RAFL, SSRs, SNPs etc. (Doğan 1997; Tunçtaner 2007; Ulaszewski et al. 2021).
For measurements and observations made on any character of organisms, phenotype emerges as the sum of genotype and environmental effects (Mettler and Gregg 1969; Ürgenç 1982; Alptekin 1986; Işık 1998). The phenotypic value of the observed character is equal to its genotypic value if the deviation caused by environmental factors is eliminated, that is, individuals are reared under the same environmental conditions in all respects (Işık 1998).
Oriental beech, which constitutes in Turkey a large part of the natural range, spreads naturally in Bulgaria, Turkey, the Caucasus and Iran in the world (Atalay 1992; Anşin and Özkan 1997). The species that is important in terms of forest volume and providing high quality timber, is one of the most economically important native deciduous tree species in Turkey (Atalay 1992; Atik 2013). Beech forests should be considered for environmental protection besides being important for timber production and should be considered as an alternative to coniferous trees (Koski and Antola 1993). It is reported that beech forests have significant differences in their habitat, thus site, yield potential and floristic composition features (Kandemir et al. 2016). Moreover, various studies have been carried out on the morphological and physiological leaf characteristics of oriental beech (Güney et al. 2016a; Bijarpasi et al. 2019; Turfan et al. 2019), leaf SPAD value and photosynthetic pigment contents (Bayraktar and Atar 2021, Bayraktar et al. 2021), seed sugar content (Güney et al. 2013), seed characteristics and germination (Vornami et al. 2004; Yılmaz 2008; Turfan et al. 2017), seed transfer regimentation (Atalay 1992), essential fatty acids, amino acids and minerals in seed (Ayaz et al. 2011), origin trials (Eyüboğlu et al. 1993; Tosun 1993), some morphological variations depending on vegetation periods (Güney et al. 2006), micro-morphological characteristics of leaf and pollen grains (Panahi et al. 2017), morphological and physiological changes related to seedlings, quality classification and land performance (Konnert and Ruetz 2003; Güney et al. 2016b; Gülseven et al. 2019; Güney et al. 2020), artificial regeneration (Tosun and Gülcan 1985) and tending intervention (Guner et al. 2017; Güney et al. 2021). However, there is no detailed study investigating the effects of geoclimatic factors on morphogenetic properties of oriental beech in Turkey.
There are two primary aims of this study:
(i) the determination of morphogenetic variations related to seed and seedling characters according to geoclimatic factors in populations that represent the natural distribution of oriental beech in Turkey;
(ii) and providing basic information about gene conservation and management of oriental beech forests by morphogenetically examining the variations among and within populations in the natural distribution of oriental beech in Turkey.
2 Material and Methods
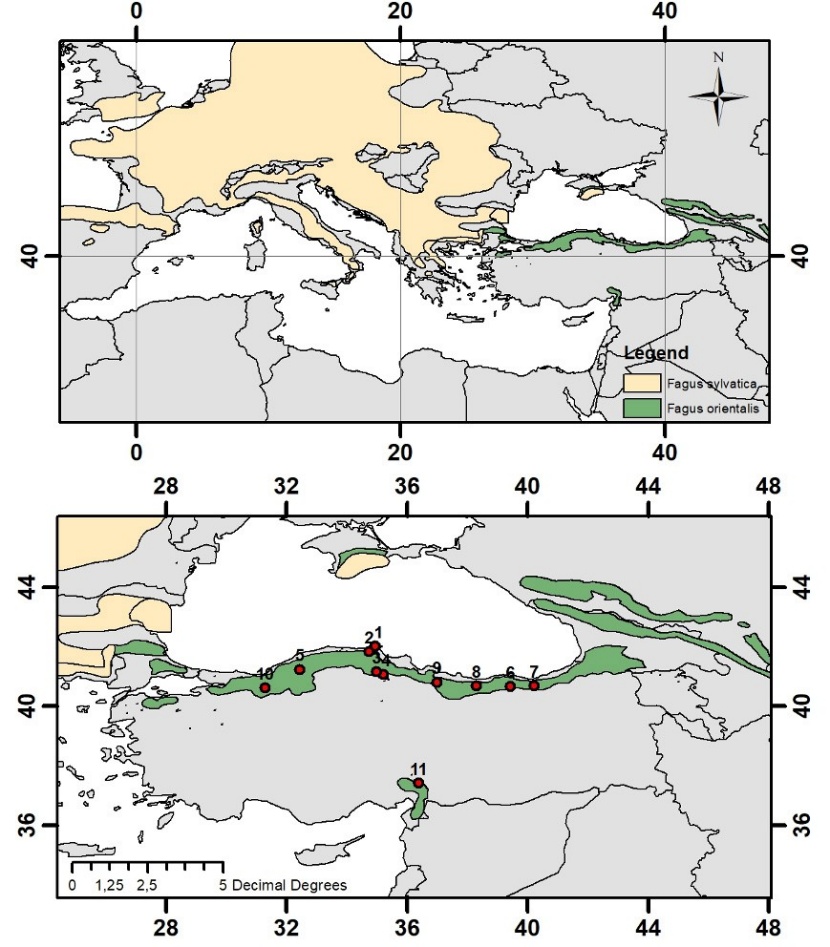
Figure 1: The distribution of sample plots in natural ranges of oriental beech in Turkey (Euforgen 2009). / Abbildung 1: Die Verteilung der beprobten Populationen im natürlichen Verbreitungsgebiet der Orientbuche in der Türkei (Euforgen 2009).
2.1 Population characteristics and sampling
In this study, 11 natural oriental beech populations, representing the natural distribution of oriental beech (Fagus orientalis Lipsky) areas in Turkey, were selected. Seeds collected from 225 trees in total, an average of 20 trees from each population, and one and two-year-old seedlings grown from these seeds were used.
Attention has been paid to the fact that the selected populations are natural, have not undergone unplanned intervention as much as possible, and especially to select populations from pure stands with normal or near-normal closure. In the selection of trees, at least 150 m distance is left between the sample trees in order to represent the population and to minimize the possibility of kinship (Vornami et al. 2004). The geographical locations of the populations where the seed materials are collected are given in Figure 1, and the information on the habitat and climate is given in Table 1.
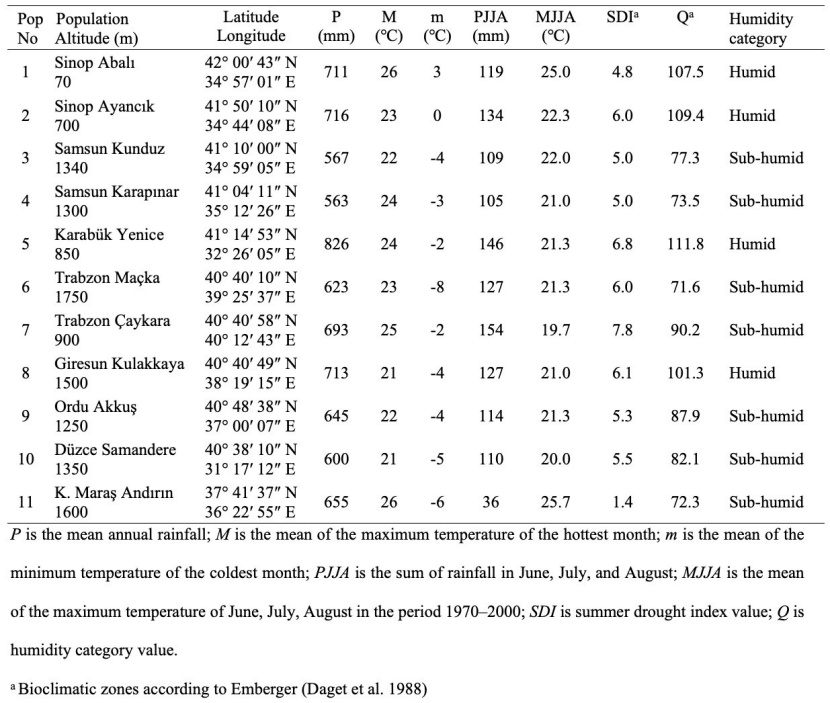
Table 1: Information about geographical and climatic characteristics of the populations. / Tabelle 1: Informationen zu geografischen und klimatischen Merkmalen der Populationen.
The climatic data needed in the study were obtained from the Worldclim database, which provides global climatic data as spatial data layers (Fick and Hijmans 2017).
Summer drought index (SDI) refers to the severity of the summer drought, as this value increases, there is a decrease in summer drought conditions. Q explains climatic humidity and temperature, the lower this value, the drier conditions occur. The coefficient “m” represents the duration of the frost cycle, higher values mean there is a shorter cycle (Balekoğlu et al. 2020). In the present study, Emberger’s equation was used to categorize the bioclimatic zones (Daget et al. 1988). The pluviothermic quotient (Q) and SDI were calculated using the equations given in Eq. 1.
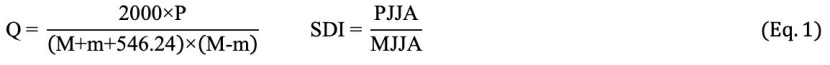
In Q equation, P is the mean annual rainfall (in mm), M is the mean of the maximum temperatures in the hottest month (in °C), and m is the mean of the minimum temperatures in the coldest month (in °C). In SDI equation, PJJA is the sum of the rainfall in June, July, and August, and MJJA is mean of the maximum temperatures in these months.
2.2 Seed and seedling material
Regarding oriental beech seeds, seed width, seed length and seed thickness were measured on the full seeds. These measurements were made with the help of digital calipers with an accuracy of 0.01 mm. In addition, 1000-seed weight was calculated according to the Eq. 2 (Ista, 1996).
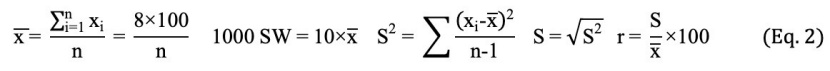
In this equation, n is the number of samples, xi is the 100-seed weight in repetitions (g), x̅ is the mean 100-seed weight, S2 is variance, S is the standard deviation, and r is the coefficient of variation.
2.3 Determination of seed vitality
Shear and tetrazolium tests were used to determine the vitality of seeds without germination. The shear tests were performed on 30 seeds from each tree. The vitality of the seeds was determined by the color and the health of the endosperm. In healthy seeds, attention has been paid to the features such as freshness of the endosperm and filling the seed coat, showing a healthy color, containing sufficient amount of water, and having a characteristic odor (Kulaç et al. 2009; Bonner and Karrfalt 2008; Güney et al. 2018). In the tetrazolium test, a solution of tetrazolium was added to cover the prepared seeds and kept in the oven at 30 °C for 10-12 hours. After 12 hours, the seeds were taken out of the oven, fixed with 70% alcohol, and the vitality of the seeds was determined according to the staining condition of the seeds (Gugala 2002).
The seeds were planted in “Of Forest Nursery” affiliated to Trabzon Regional Directorate of Forestry. The measurements in the seedlings were made separately after the completion of two growth periods. For this, seedling length, root collar diameter and number of side branches were measured separately in a total of 6750 seedlings to be 30 seedlings for each tree in 225 trees belonging to 11 populations. Seedling length was taken as the length between the tip of the terminal bud from the root collar and measured at 0.1 cm sensitivity with the help of a ruler. The root collar diameter of each seedling was determined by measuring with the help of a digital caliper with a sensitivity of 0.01 mm. The number of side branches per seedling was determined by counting the side branches on the seedling one by one. The list of measured characters of seed and seedlings with measurement and units are given in Table 2.
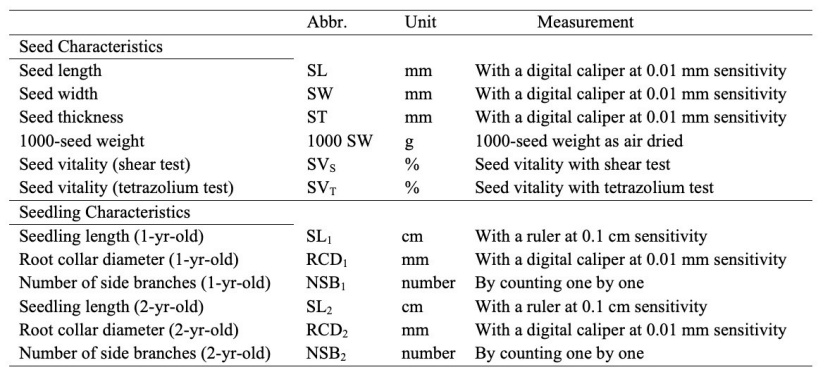
Table 2: Morphometric characters measured for seeds and seedlings. / Tabelle 2: Morphometrische Merkmale, die bei Samen und Sämlingem gemessen wurden.
2.4 Statistical Analysis
The obtained data was analyzed using SPSS 20 and R statistical packages. Before the calculations, data was standardized and the morphological distance among seed stands were estimated. Differences among the populations were determined using the analysis of variance (one-way ANOVA). In order to determine the groups occurring among the populations, Duncan's test was used, and to measure the distances among the populations hierarchical cluster analysis was used. Factor analysis was carried out to determine the possibilities of revealing the variation determined among and within the populations, by using fewer variables. Statistically significant models were determined by performing regression analysis to estimate seedling characters. Relationships among the morphological characteristics were tested using the correlation analyses. Moreover, the data were analyzed using Penrose's analysis (Eq. 3 and 4) to show the differences among populations mathematically.

In Eq. 1, Zi,k is standardized value of the kth characteristics of the ith population, Xi,k refers to mean of the kth characteristics of the ith populations for the kth characteristics, and Sk stands for the standard deviation of the populations for the kth characteristics (Şevik et al. 2010).

In Eq. 2, Di,j means the morphological distance between the ith population and the jth population, n refers to the number of characteristics, μkj stands for the standardized values of the kth characteristics of the jth population, and Vk is the variance of standardized averages of the kth characteristics (Şevik et al. 2010).
3 Results
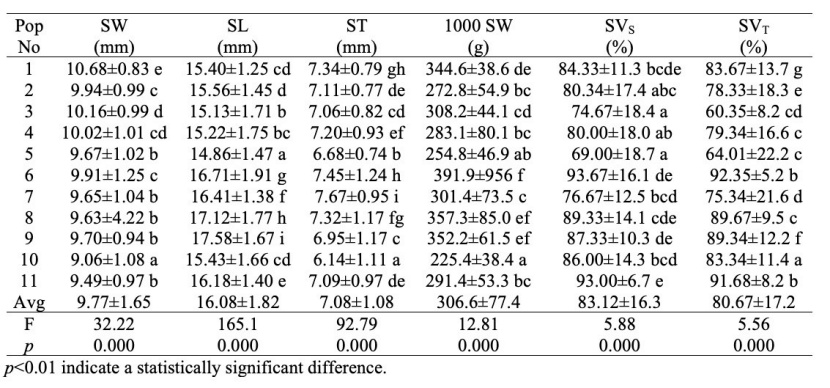
The averages of the seed characteristics of the 11 populations subject to the study and the results of the analysis of variance (one-way ANOVA) and the groups formed as a result of Duncan's test are given in Table 3.
We found statistical differences among the populations depending on the data on seed characteristics measured in the study. The averages of SW, SL and ST for all populations were determined as 9.77 mm, 16.08 mm and 7.08 mm, respectively. As a result of Duncan's test regarding 1000-seed weight in F. orientalis, eight different groups were formed. Accordingly, while Trabzon-Maçka population had the highest 1000-seed weight with 391.91 g, the Düzce-Samandere population had the lowest 1000-seed weight with 225.38 g, and the average 1000-seed weight for all populations was determined as 307.54 g.
As a result of the analysis of variance related to the seed shear test, it was determined that the seed vitality rates differ according to the populations and nine different groups were formed as a result of Duncan's test. As a result of the shear test, the Kahramanmaraş-Andırın population had the highest seed vitality percentage with 93%, while the Karabük-Yenice population had the lowest seed vitality percentage with 69%.
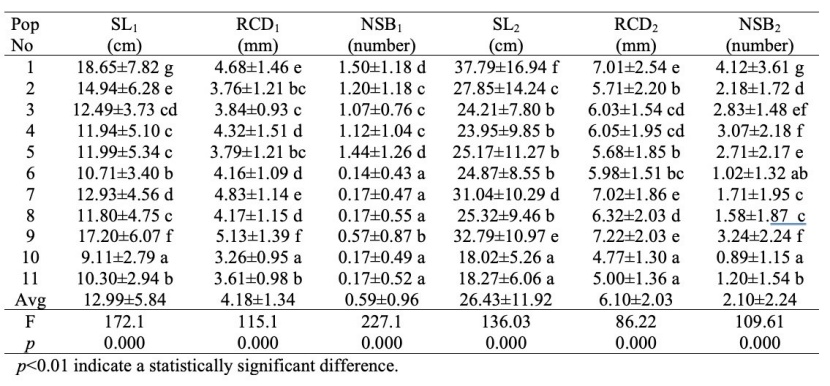
Table 4: Results seedling characteristics, variance analysis and Duncan's test. For explanation of abbreviations please see Table 2. / Tabelle 4: Ergebnisse der Merkmale der Sämlinge, der Varianzanalyse und des Duncan-Tests. Für Erklärung der Abkürzungen siehe Tabelle 2.
It was determined that the populations showed variation in terms of seed vitality rates related to the tetrazolium test and five different groups occurred as a result of the Duncan's test. According to the tetrazolium test, the Trabzon-Maçka population had the highest seed vitality percentage with 96.7%, while the Samsun-Kunduz population had the lowest seed vitality percentage with 60.35%. Although the percentage of vitality determined as a result of the shear test is parallel to the results of the tetrazolium test, generally the percentage of vitality determined by the shear test was higher.
The average values for the seedlings, the results of variance analysis and the groups formed as a result of the Duncan's test are given in Table 4.
There were statistical differences among the populations regarding all the measured seedling characteristics, and Table 4 also shows how the seedlings of one and two-year-old are grouped according to the Duncan’s test. Sinop-Abalı and Ordu-Akkuş populations, where the highest average seedling length was observed, were again in the first place in terms of seedling length at the end of the second vegetation period. However, the populations of Düzce-Samandere and Kahramanmaraş-Andırın, which had the lowest average at the end of the first vegetation period in terms of seedling length, also had the lowest average when they reached the age of 2. In terms of root collar diameter, Ordu-Akkuş, Sinop-Abalı and Trabzon-Çaykara were the populations with the highest average, Düzce-Samandere and Kahramanmaraş-Andırın were the populations with the lowest average, and this situation was the same at the end of both the first and the second vegetation period.
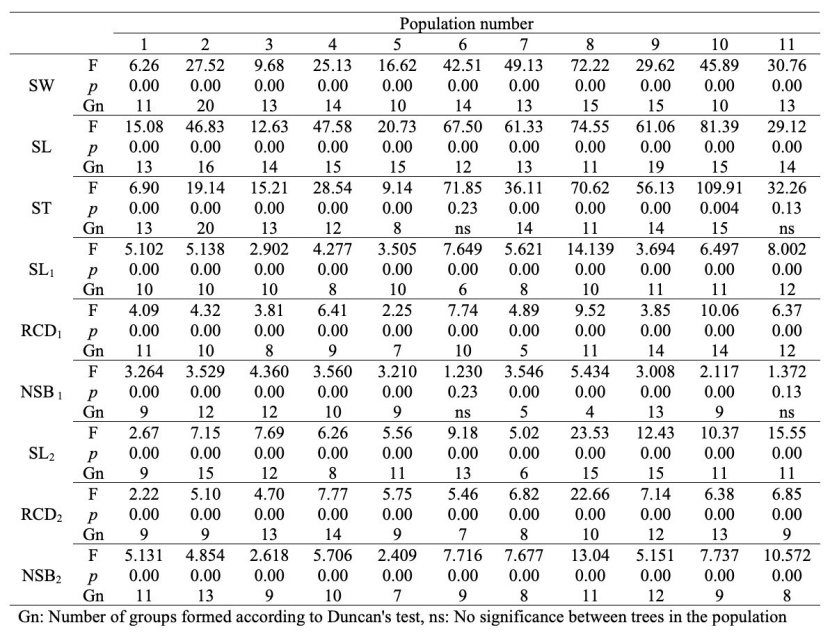
Table 5: Variance analysis results for seed and seedlings within populations. For abbreviations, see Table 2. / Tabelle 5: Ergebnisse der Varianzanalyse für Samen und Sämlinge innerhalb von Populationen. Für Erklärung der Abkürzungen siehe Tabelle 2.
Seed and seedling-related characters were also determined separately in trees within the populations, and the results of variance analysis revealing whether these values differ within populations are given below (Table 5).
When Table 5 is examined, it was determined that the populations of Trabzon-Maçka and Kahramanmaraş-Andırın did not differ in terms of ST and NSB1, while the other populations showed variation in terms of all characters and formed many different groups.
When calculating morphological distances, different characteristics with different units are brought together, the Penrose equation is obtained. As the result of the equation approaches zero, the similarity among populations increases. All values measured for the 11 populations subject to the study were included in Penrose's analysis (Pn) and the morphological distances of the populations from each other were revealed (Table 6).
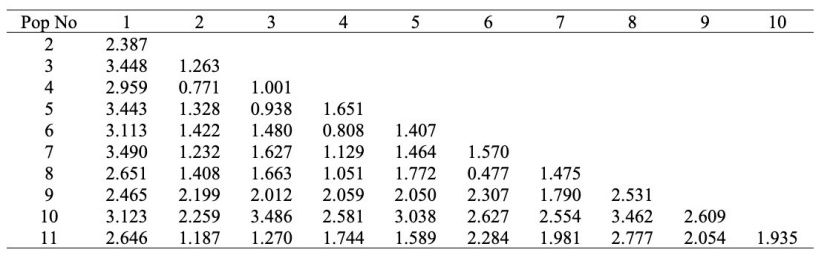
Table 6: Morphological distance values among populations using Penrose's analysis. / Tabelle 6: Morphologische Distanzwerte zwischen Populationen auf Basis von Penrose Analyse.
As can be seen from Table 6, the highest distance value was found between the Sinop- Abalı and Trabzon-Çaykara populations (Pn1,7: 3.490) and the lowest distance value was found between the Trabzon-Maçka and Giresun-Kulakkaya populations (Pn6,8: 0.477).
Therefore, according to Penrose's analysis results, the Sinop-Abalı and Trabzon-Çaykara populations were determined as the most distant populations, while the Trabzon-Maçka and Giresun-Kulakkaya populations were the closest to each other.
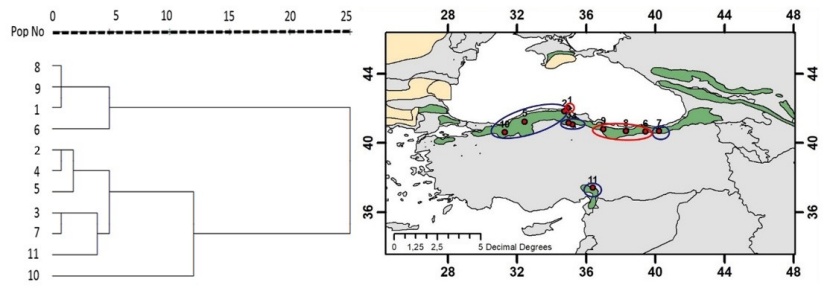
Figure 2: Dendrogram for cluster analysis and representation of the formed groups in the study region. / Abbildung 2: Dendrogramm zur Clusteranalyse und Darstellung der gebildeten Gruppen im Untersuchungsgebiet.
The groups formed by the populations as a result of cluster analysis regarding all measured characters and the distribution of these groups on the map are given in Figure 2.
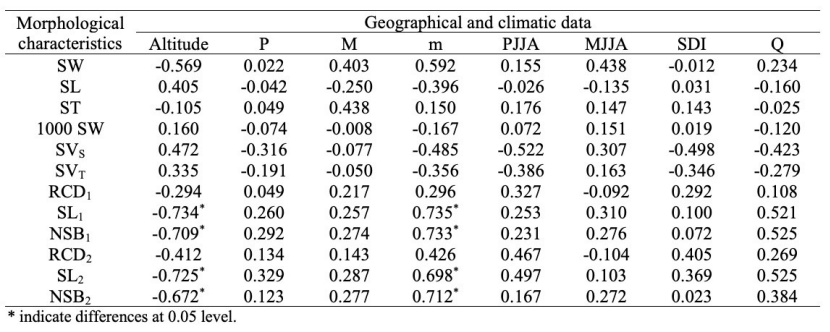
Table 7: Correlation coefficients of seed, seedling, altitude, and climatic variables. For abbreviations, please see Tables 1 and 2. / Tabelle 7: Korrelationskoeffizienten von Samen, Sämling, Seehöhe und Klimavariablen. Für Abkürzungen siehe Tabellen 1 und 2.
The significance of the groupings made by cluster analysis was tested with the discriminant analysis and as a result, it was found significant to divide the populations into 2 groups. As can be seen from Figure 2, the Sinop-Abalı, Trabzon-Maçka, Giresun-Kulakkaya and Ordu-Akkuş populations are in one group, while the other populations constitute the second group. As a result of Penrose's analysis, the Sinop-Abalı and Trabzon-Çaykara populations, which have the greatest genetic distance, were also in a separate group in the dendrogram formed as a result of cluster analysis, while the Trabzon-Maçka and Giresun-Kulakkaya populations, which have the least genetic distance, were in the same group in the dendrogram formed as a result of cluster analysis (Table 6, Figure 2). The results of the correlation analysis made between the characters measured for the populations in our study and the climate data and bioclimatic zones are given in Table 7.
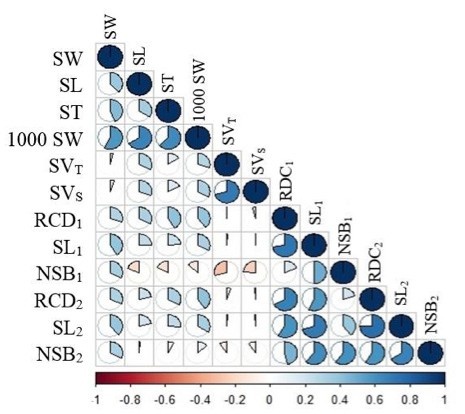
Figure 3: Graphic representation of the correlation analysis. / Abbildung 3: Graphische Darstellung der Korrelationsanalyse.
As a result of the correlation analysis between bioclimatic zones and seed and seedling morphological characteristics, positive correlations were found between m (mean of the minima of the coldest month) and SL1, NSB1, SL2 and NSB2, while negative correlations were found between altitude and these characteristics (Table 7). According to these data, it is seen that seedling length and root collar diameter values in seedlings grown from the seeds obtained from the populations increase, as the mean of the minima of the coldest month value in the long-term increases and the altitude decreases. The direction and strength of the correlation between the data of the seed and seedling characteristics of the populations are schematized in Figure 3.
According to the results of the correlation analysis between the measured characters, the seed sizes show statistically significant correlations with each other and with SL1-2, RCD1-2 and NSB1-2 values of the one and two-year-old seedlings. It is understood that the shear test and the tetrazolium test performed to determine the seed vitality show a significant correlation. In addition, a positive correlation was found between the results of the tetrazolium test and seed length. Moreover, a positive correlation between 1000-seed weight and SL1-2 and RCD1-2 values of the one and two-year-old seedlings reveals that this character is related to the seedling growth.
Based on the one-year-old seedling characters of all populations, statistically significant models were determined by performing regression analysis to estimate the two-year-old seedling characters. Accordingly, it was determined that a linear pattern was formed between the seedling length of two-year-old and the seedling length and root collar diameter of one-year-old seedlings, and a quadratic pattern with the number of side branches. The equation (Eq. 5) for the seedling length of the two-year-old seedling is given below.
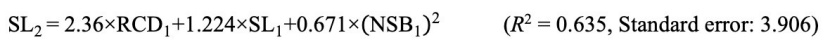
As a result of the regression analysis performed to estimate the root collar diameter of two-year old seedlings with morphological measurements of one-year old seedlings, it was determined that the linear pattern was statistically significant between the root collar diameter of two-year-old seedlings and the seedling length and root collar diameter of the one-year-old seedlings. Accordingly, the equation (Eq. 6) that can be used to estimate the root collar diameter of two-year-old seedling is as follows.
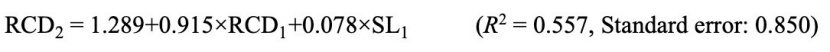
Factor analysis was carried out to determine the possibilities of revealing the variation determined among and within the populations depending on the measured characters related to seed and one and two-year-old seedlings, by using fewer variables. In order to better reveal the relationships between variables, rotation was made according to the Varimax method. For this purpose, factor analysis was performed with variables related to SL, SW, ST, 1000 SW, SVS, SVT, SL1, RCD1, NSB1, SL2, RCD2 and NSB2. As a result of the factor analysis, it was determined that instead of these 12 different variables, using 4 variables including SL1, RCD1, SL2 and RCD2, the variation among and within populations could be revealed statistically with an explanatory rate of 68.9%.
4 Discussion
4.1 Seed characteristics
In this study, the variations of the seed and seedling characteristics of the 11 oriental beech (Fagus orientalis Lipsky) populations were determined related to the populations and the relations of these variations with the climate data were revealed. According to the results of the study, various variations were determined in terms of seed sizes among and within populations on tree basis. In the study conducted in Fagus sylvatica L. by Tylek and Walczyk (2002), the average seed width was determined as 8.8 mm, seed length as 16.93 mm and seed thickness as 7.20 mm. As can be seen from this study, it can be said that F. sylvatica and F. orientalis have similar values in terms of seed sizes. There are many studies that determine the differences among and within populations regarding seed sizes. Depending on the seed characteristics, in the studies performed in F. sylvatica (Bodyl 2007) and Fagus crenata Blume (Hiura et al. 1996), differences among the populations were determined in terms of seed sizes. In another study performed in F. sylvatica, while determining the variations regarding seed sizes, it was reported that seed sizes had positive effects on germination and seedling development (Thomsen and Kjaer 2002).
In various studies carried out in F. orientalis, it was determined as an average of 273.1 g (215.3–316.7 g) by Saatçioğlu and Ürgenç (1960), an average of 292.2 g by Tosun (1993) and an average of 307.3 g Yılmaz (2005). In F. sylvatica, 1000-seed weight was determined as 249.2 g by Tylek and Walczyk (2002) and 209.8 g by Suszka and Kluczynska (1980). Yılmaz (2005) found the vitality rate as 86.19% as a result of the shear test performed on 14 origins in his study on F. orientalis. These values are consistent with the results we obtained with the F. orientalis. Additionally, in a study conducted on F. orientalis to determine seed vitality by shear test in various origins, seed vitality percentage was determined as 49.83% (Gugala 2002).
In a study conducted in F. orientalis, the vitality rate of 14 origins was found to be 79.43% as a result of the tetrazolium test, while significant relationships were found between tetrazolium test and shear test (Yılmaz 2005). In one research by Gugala (2002) performed in F. sylvatica, the vitality percentage according to the tetrazolium test was determined as 88.25% and the average germination percentage was 76.25%. In another study comparing the results of the tetrazolium test and germination test, tetrazolium test and germination percentage values were found to be 50.07% and 42.41% for Abies nordmanniana Spach, and 52.54% and 43.36% for Cedrus libani A.Rich., respectively (Velioğlu and Arslan 2000). Therefore, it is stated that there is a high correlation between the results of both tests.
4.2 Seedling characteristics
In this study, it was aimed to reveal the genetic differences depending on morphological characteristics related to the seedlings by keeping the environmental factors homogeneous in nursery. As a matter of fact, in similar studies, it is reported that the morphological characteristics determined in this way reflect genetic characteristics. In an investigation into the relationship between morphological features and genetic features in Raphanus sativus L., genetic characteristics were compared with the morphological characteristics of germination, seed and seedling and RAPD method and as a result, significant relationships between genetic and morphological characteristics were determined (Pradhan 2004). The highest values in terms of seedling length and root collar diameter in Sinop-Abalı population may be due to the very low altitude in this population compared to others. Some studies in F. orientalis have revealed that altitude is also an important factor in genetic variation. The study investigating genetic variations in F. sylvatica populations between 150-600 m altitudes with the method of starch gel electrophoresis by Sander et al. (2000) can be given as example to this situation. As a result of their study, significant differences were determined between populations, while the highest difference was found in the population with the highest altitude.
The spread of F. orientalis in a very wide geographical area in Turkey and the difference in ecological factors both among and within the population in these geographical regions can be explained as the reason for the genetic variations in this study. In many studies performed in F. sylvatica, the reason for the genetic differentiation in beech stands is explained as natural selection and genetic isolation together with the differences in environmental factors due to the wide geographic spread of the species (Barrière et al. 1985; Cuguen et al. 1985; Belletti et al. 1996), and this explanation supports our results.
Since forest trees can transport most of their seeds to a narrow area, each population consists of subpopulations that make up them (Işık 1988). Since the subpopulation consists of different individuals which is a result of adaptation to local environmental differences under the influence of different selection pressure and migration factors special to the specific environmental conditions of the microhabitat, the genetic diversity within the population is high (Işık 1983). Therefore, different races and sub-races can occur even over very short distances. There are studies revealing the existence of different local forms over short distances in Turkey (Boydak 1977). This situation is similar to the F. orientalis discussed in the study. As it is known, beech seeds are coarse and can germinate under their own cover, that is, they can create micro-environments in very narrow areas. Conserving the genetic diversity of a species has paramount importance for climate change adaptation. The loss of genetic diversity is probably because of the narrowing of the species' range (Alsos et al. 2012). Due to the wide distribution loss, especially the oriental beech may experience a loss of genetic diversity (Yalcin, 2012). For this reason, current marginal populations are important tools for maintaining genetic diversity in the future (Barbati et al. 2018).
In the present study, when the differences between the average values of the populations and the differences between the trees within the populations are examined, it is understood that the differences in the seedling-related characters within the populations are more than the difference among the populations. There are many studies in which genetic variations are determined among and within populations regarding the morphological characteristics of seeds, saplings, needles and leaves, DNA and isoenzyme analyzes in different beech species. In the studies performed in F. sylvatica regarding different climate, soil and topographic features (Comps et al. 1991), different altitude zones (Sander et al. 2000), DNA diversity in the chloroplast (Vettori 2004; Magri et al. 2006), isoenzyme analysis on individuals in the population (Krzakowa and Matras 2005), polymorphic enzyme systems and different enzyme systems (Merzeau et al. 1994; Comps 1998), variations among and within the populations were revealed. Genetic differences were determined in different studies that isoenzyme analyzes were performed in F. orientalis and F. sylvatica (Pastorelli et al. 2003), and conducted by sampling 14 populations regarding DNA diversity in chloroplast in F. orientalis (Shanjani et al. 2004). In other studies, conducted in F. crenata, differences were determined in 11 enzyme systems related to DNA in chloroplast and performed by sampling buds (Tomaru et al. 1998; Fujii et al. 2002). In the study carried out in Fagus moesiaca (K.Malý) Czeczott, differences among populations with isoenzyme analyzes were revealed (Gömöry et al. 1999). In the study carried out by DNA isolation using leaves to determine the species difference in F. orientalis and F. sylvatica populations naturally spread in Turkey, no different grouping was observed between F. orientalis and F. sylvatica (Kandemir et al. 2016).
On the other hand, studies reveal that forestry practices and silvicultural interventions in forest affect genetic diversity. In a study conducted on Picea glauca (Moench) Voss, it was determined that stands performed natural regeneration and natural aged pure stands in terms of general average have higher genetic diversity compared to artificially established plantations without breeding work. Moreover, it was stated that the low genetic diversity in plantations is due to regeneration with individuals whose genetic basis is not wide (Rajora 1999). In another study conducted in F. crenata, it was determined that the cuttings made in the forests caused changes in genetic diversity (Takahashi et al. 2000).
The high genetic diversity determined within the populations in the present study is of great importance in terms of preserving this genetic base in the population and avoiding interventions that may cause genetic pollution. As a matter of fact, it has great importance that the genetic base in a population is formed over very long periods and accordingly, individuals with the highest adaptation value to that region are in the population. If this wealth is not preserved or if individuals with unknown genetic basis are brought to the area, the existing genetic wealth may be destroyed (Hamrick 1985). There are various studies that reveal this situation. Yalcin (2012) in a study on the natural distribution area of eastern beech in Turkey reported that most of the available suitable areas will disappear at the end of 2050, and only 3-7% of the existing stands will remain suitable until 2080 with optimistic estimates and the change in suitable areas will be dramatic. Furthermore, Ayan et al. (2022) and Dagtekin et al. (2019), stated that by 2070, the oriental beech will greatly decrease in the south and southwest regions of Turkey and will completely lose its isolated marginal populations in the Amanos Mountains and the species will continue to exist especially in the Caucasus.
As a result of a study conducted on F. sylvatica in Italy, it was determined that only 4.3% of the total genetic variation was caused by differences among populations (Belletti et al. 1996). Similar results were obtained in other studies performed in F. sylvatica, and the differences among populations were found to be low (Comps et al. 1991; Konnert 1995). In a similar study performed in 16 populations in F. japonica, it was stated that the variation among population was low and the variation within the population was high (Hiraoka and Tomaru 2009). As a result of gene flow, obtained offspring without fertilization among populations may be less adaptive than obtained offspring without fertilization within populations (Millar and Libby 1989). Outbreeding depression can lead to various adjustment disorders and even deaths in the new generation. As a result of the contamination of the gene pool of the natural population, many gene combinations adapted to the local conditions are dispersed. For this reason, breeding studies and genetic planning should be done especially in order to ensure the continuity of local forms and as a precaution against genetic pollution (Velioğlu et al. 2004).
It is stated that the genetic structure in beech stands depends on natural selection, except for gene flow and genetic drift. In addition, various studies have reported that these factors cause differences among and within populations (Müller-Starck 1985). Previous researches have stated that the genetic variation in wind-pollinated species such as beech is high (Hamrick et al. 1992) and the variation within the population is generally higher, and the variation among populations generally does not exceed 5% (Larsen 1996).
4.3 Morphogenetic distance between populations
According to the results of Penrose's analysis performed to determine the genetic distance between populations, the genetic distance value of the Samsun-Kunduz and Samsun-Karapınar populations, which are closer to each other in terms of geographic distance, is Pn3,4: 1.001, revealing that these populations are close to each other in terms of genetic distance. However, in the Trabzon-Maçka and Trabzon-Çaykara populations, which are relatively close in terms of geographical distance, the fact that the distance value is Pn6,7: 1.570 indicates that the genetic distances are not very close. Again, the distance value Pn2,11: 1.187 in the Kahramanmaraş-Andırın and Sinop-Ayancık populations, which are very distant from each other in terms of geographic distance, indicates that these populations are genetically relatively close. As can be understood from these results, there is no direct relationship between the geographical distance of the populations to each other and their genetic distance from each other.
Various studies have been conducted to examine the relationship between geographic distances and the genetic characteristics of populations. In different studies carried out in F. sylvatica, it was stated that there was no significant difference between the genetic distances of the populations depending on the geographical distances (Belletti et al. 1996; Gömör et al. 2003). In a similar study conducted in F. sylvatica, it was stated that there are similarity trends in the genetic structure of trees with distances fewer than 30 m from each other, but variations in the genetic structure are affected by different factors depending on the geographical distances (Vornami et al. 2004).
Considering that the natural distribution areas of the oriental beech in Turkey are very variable in terms of features such as topographic structure and climate, and it varies even in short distances, it can be said that there is no direct relationship between geographical distances and genetic distances for this species. As a matter of fact, according to Emberger's bioclimate zones (Daget et al. 1988), the Sinop-Abalı, Sinop-Ayancık, Karabük-Yenice and Giresun-Kulakkaya populations are included in the humid section in the humidity category, while other populations are in the sub-humid section.
The high genetic diversity among and within population detected in all characters observed in seeds and seedlings in our study continued in the characteristics of the seedlings studied at the end of the first and second vegetation periods. According to the results obtained in terms of seed and seedling characteristics in oriental beech, it was seen that the genetic diversity was quite high between families among and within the populations. In the selection of seed sources in order to grow quality seedlings, first of all, suitable population or origins, then suitable families or genotypes in these populations or origins should be determined with great care.
5 Conclusions
As a result of this study, which was carried out to determine the morphogenetic variations among and within population of the oriental beech, it was revealed to show variations among and within population both within the optimal distribution areas and in the natural populations outside this area. Considering the variations within the selected populations in the study, it was determined that there are more differences in characteristics within the populations compared to the characteristics among the populations. Therefore, the source of the variation seems to be within populations rather than among populations.
According to the results of the study, the Sinop-Abalı, Trabzon-Maçka, Giresun- Kulakkaya and Ordu-Akkuş populations were separated from the other populations by generally taking part in the same group in terms of characters measured in 11 different populations. However, among these populations, the Sinop-Ayancık population and especially the Sinop-Abalı population are outside the optimal natural range of oriental beech in terms of altitude and show variations markedly. Therefore, conservation of gene resources in situ is important in terms of continuity of variation and thus conservation of biological diversity. For this, it is necessary to protect the existing populations and ensure the continuity of genetic diversity considering the significant risks in the natural distribution of oriental beech, especially according to long-term climate scenarios.
According to the groups obtained in the present study, showing typical distinctions and taking place in different groups; separating the populations such as Sinop-Abalı, Sinop-Ayancık, Kahramanmaraş-Andırın, Trabzon-Maçka and Düzce-Samandere in terms of seed transfer and using only the seed and seedling material obtained from the populations in the forest establishment studies to be conducted in these regions, both to protect the determined genetic diversity and to prevent genetic pollution may be important.
When classifying seeds in oriental beech, instead of using homogeneous groups in terms of seed sizes by classifying them according to certain sizes, it will be more beneficial to use seeds of all sizes and quality that increase the variety. However, thanks to the regression analysis created based on the data obtained in the study, it can be estimated the morphological values of two-year-old seedlings with the morphological data of one-year-old seedlings, and thus this knowledge can be used both in breeding studies and in determining the seedling growth course.
In forest establishment studies to be carried out in oriental beech, attention should be paid to both altitude zones and preservation of variation value. With the use of seedlings to be grown by using seeds of a certain origin in these studies, success will increase and genetic losses will be eliminated by using suitable origins.
Acknowledgements
We would like to thank Alkan Günlü for his great supports in collecting the seeds. We also thank Trabzon-Of Forest Nursery staff for their support.
Funding
This work was financially supported by The Scientific and Technological Research Council of Turkey (Project No: 105O534) and Research Fund of the Karadeniz Technical University (Project No: BTAP-360).
Conflict of Interest
The authors declare that they have no conflict of interest.
References
Alan M (2017) Importance of marginal populations for conservation of forest gene resources. Biological Diversity and Conservation 10/2(S2): 55-62.
Alsos IG, Ehrich D, Thuiller W, Eidesen PB, Tribsch A, Schönswetter P, Lagaye C, Taberle TP, Brochmann C (2012) Genetic consequences of climate change for northern plants. Proc. R. Soc B: Biological Sciences, 279(1735):2042-2051. https://doi.org/10.1098/rspb. 2011. 2363.
Anşin R, Özkan ZC (1997) Tohumlu bitkiler, odunsu taksonlar. KTÜ Orman Fakültesi Yayın No. 19, Trabzon.
Atalay İ (1992) Kayın (Fagus orientalis Lipsky) ormanlarının ekolojisi ve tohum transferi yönünden bölgelere ayrılması. Orman Bakanlığı, Orman Ağaçları ve Tohumları Islah Araştırma Müdürlüğü, Yayın No. 5, Ankara, p 209.
Atar F (2021) Effects of altitude on some seedling quality characteristics of Carpinus betulus L. (Common Hornbeam) and Carpinus orientalis Mill. (Oriental Hornbeam). Artvin Çoruh Üniversitesi Orman Fakültesi Dergisi, 22(2): 257-265.
Atar F, Bayraktar A, Atar E, Turna İ (2017) Morphological Diversity of Common Hornbeam (Carpinus betulus L.) Seeds in the Eastern Black Sea Region of Turkey. International Forestry and Environment Symposium, Trabzon, Turkey, 7 - 10 November 2017.
Atar F, Bayraktar A, Yıldırım N, Turna İ, Güney D (2020) Fruit and seed diversity of Smilax excelsa in the Black Sea Region, Turkey. Ormancılık Araştırma Dergisi, 7(1): 1-8.
Atar F, Güney D (2021) Doğu Gürgeni (Carpinus orientalis Mill.) Fidanlarında Farklı Önişlem ve Yükseltiye Bağlı Olarak Klorofil İçeriğinin Değişimi. Bartın Orman Fakültesi Dergisi, 23(1): 236-243.
Atar F, Turna I (2018) Fruit and seedling diversity among sweet chestnut (Castanea sativa Mill.) populations in Turkey. Šumarski list, 142(11-12): 611-619.
Atik A (2013) Effects of planting density and treatment with vermicompost on the morphological characteristics of oriental beech (Fagus orientalis Lipsky). Compost Science and Utilization 21: 87-98. https://doi.org/10.1080/1065657X.2013.836066
Ayan S, Bugday E, Varol T, Özel HB, Thurm EA (2022) Effect of climate change on potential distribution of oriental beech (Fagus orientalis Lipsky.) in the twenty-first century in Turkey. Theor Appl Climatol 1-13. https://doi.org/10.1007/s00704-022-03940-w
Ayaz FA, Glew RH, Turna İ, Güney D, Chuang LT, Chang YC, Andrews R, Power L, Presley J, Torun H, Sahin N (2011) Fagus orientalis (oriental beechnut) seeds are a good source of essential fatty acids, amino acids and minerals. Food 5(SI1): 48-51.
Balekoglu S, Caliskan S, Dirik H (2020) Effects of geoclimatic factors on the variability in Pinus pinea cone, seed, and seedling traits in Turkey native habitats. Ecological Processes 9: 55. https://doi.org/10.1186/s13717-020-00264-3
Barbati A, Scarascia G, Ayan S, Blasi E, Calama R, Canaveira P, Cicatiello C, Collalti A, Corona P, Del Rio M, Ducci F, Perugini L (2018) State of Mediterranean forests 2018. Chapter 8: Adaptation and mitigation, p. 128–146, Published by the FAO of the United Nations and Plan Bleu, Regional Activity Center of UN Environment / Mediterranean Action Plan, ISBN FAO: 978–92–5–131047–2 ISBN Plan Bleu: 978–2–912081–52–0, Rome.
Barrière G, Comps B, Cuguen J, Nitsiba F, Thiebaut B (1985) The genetical ecological variability of beech (Fagus sylvatica L.) in Europe - an alloenzymatic study: genetic isolations of beechwoods. In: Proc 1st Symp Improvement and Silviculture of Beech. IUFRO Project Group P1 10-00, Grosshansdorf, pp 24-50.
Bayraktar A, Atar F (2021) Effects of different nanotechnological fertilizer applications on SPAD value in Fagus orientalis leaves. Paper presented at 4th International Eurasian Conference on Biological and Chemical Sciences, Ankara, 24-26 November.
Bayraktar A, Güney D, Atar F (2021) Farklı nanoteknolojik sıvı organomineral gübre dozlarının doğu kayını fidanlarının fotosentetik pigment içerikleri üzerine etkileri. In: Cengizler İ, Duman S (eds) Ziraat, orman ve su ürünlerinde araştırma ve değerlendirmeler – I. Gece Kitaplığı, Ankara, pp 267-284.
Belletti P, Lanteri S (1996) Allozyme variation among European beech (Fagus sylvatica L.) stands in Piedmont, North-Western Italy. Silvae Genetica 45(1): 33-36.
Bijarpasi MM, Shahraji TR, Lahiji HS (2019) Genetic variability and heritability of some morphological and physiological traits in Fagus orientalis Lipsky along an elevation gradient in Hyrcanian forests. Folia Oecologica 46(1): 45-53. https://doi.org/10.2478/foecol-2019-0007
Bilgen BB, Güney D, Kaya N (2011) Estimation of altitudinal genetic variation in Pinus sylvestris L. populations in Trabzon by RAPD markers. Paper presented at VI. International Symposium on Ecology and Environmental Problems, Antalya, 17-20 November.
Bodyl M, Sulkowska M (2007) Estimation of European beech (Fagus sylvatica L.) seed differentiation in Poland in the years 1992-2004. Sylwan 151(9): 12-21.
Bonner TB, Karrfalt RP (2008) Seed testing. The woody plant seed manual. United States Department of Agriculture, Forest Service, Agriculture Handbook 727.
Boydak M (1977) Sarıçam (Pinus sylvestris L.) doğal populasyonlarında dikey yönde polen hareketleri ve uygulamadaki önemi. İÜ Orman Fakültesi Dergisi 27(2): 226-238.
Chmura DJ, Rozkowski R (2002) Variability of beech provenances in spring and autumn phenology. Silvae Genetica 51: 2-3.
Comps B, Matyas C, Letouzey L, Geburek T (1998) Genetic variation in beech populations (Fagus sylvatica L.) along the alpine chain and in the Hungarian basin. Forest Genetics 5(1): 1-9.
Comps B, Thiebaut B, Sugar I, Trinajstic I, Plazibat M (1991) Genetic variation of the Croatian beech stands (Fagus sylvatica L): Spatial differentiation in connection with the environment. Ann Sci For 48: 15-28.
Cuguen J, Thiebaut B, Nitsiba F, Barrière G (1985) Enzymatic variability of beech stands (Fagus sylvatica L.) on three scales in Europe: evolutionary mechanisms. In: Jacquart P, Heim G, Antonovics J (eds) Genetic Differentiation and Dispersal in Plants. NATO ASI Series, Montpellier, pp 17-39.
Daget P, Ahdali L, David P (1988) Mediterranean bioclimate and its variation in the Palaearctic region. In: Specht RL (ed) Mediterranean-type ecosystems, a data source book. Kluwer Academic Publishers, Dordrecht, pp 139-148.
Dagtekin D, Sahan EA, Denk T, Kose N, Dalfes HN (2019) Predicting late quaternary distribution and future refugia of oriental beech (Fagus orientalis Lipsky) in Asia minor. International Conference on Climate Change & Forestry, TOD Publication Number: 49, Conference Proceeding, 177-182, Antalya.
Dagtekin D, Şahan EA, Denk T, Köse N, Dalfes HN (2020) Past, present and future distributions of oriental beech (Fagus orientalis) under climate change projections. PloS ONE 15(11): e0242280. https://doi.org/10.1371/journal.pone.0242280
Doğan B (1997) Kazdağı yöresi doğal kızılçam populasyonlarında izoenzim çeşitliliği. Ege Ormancılık Araştırma Enstitüsü, 10, İzmir, 3-5.
Euforgen (2009) Distribution map of oriental beech (Fagus orientalis), www.euforgen.org.
Eyüboğlu AK, Atasoy H, Küçük M (1993) Doğu Karadeniz Bölgesinde doğu kayını (Fagus orientalis Lipsky) orijin denemelerinin dokuz yıllık sonuçları. Ormancılık Araştırma Enstitüsü Yayınları, Teknik Bülten No. 237, Ankara.
Fick SE, Hijmans RJ (2017) WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37(12): 4302-4315.
Fujii N, Tomaru N, Okuyama K, Koike T, Mikami T, Ueda K (2002) Chloroplast DNA phylogeography of Fagus crenata (Fagaceae) in Japan. Plant Syst Evol 232: 21-33.
Gömöry D, Paule L, Brus R, Zhelev P, Tomovic Z, Gracanş J (1999) Genetic differentiation and phylogeny of beech on the Balkan peninsula. J Evol Biol 12: 746-754.
Gömöry D, Paule L, Shvadchak IM, Popescu F, Sulkowska M, Hynek V, Longauer R (2003) Spatial patterns of the genetic differentiation in European beech (Fagus sylvatica L.) at allozyme loci in the Carpathians and the Adjacent Regions. Silvae Genetica 52(2): 78-83.
Gugala A (2002) Changes in quality of beech (Fagus sylvatica L.) seeds stored at the Forest Gene Bank Kostrzyca. Dendrobiology 47: 33-38.
Guner S, Tufekcioglu A, Celik N (2017a) Effects of Thinning, Liming, and Nitrogen Application on The Growth of A Young Oriental Beech (Fagus orientalis Lipsky) Forest Stand. Fresenius Environmental Bulletin 26(11): 6727-6733.
Gülseven O, Ayan S, Özel HB, Yer EN (2019) Farklı doğu kayını (Fagus orientalis Lipsky) populasyonlarına ait fidanların morfolojik ve fizyolojik karakteristikleri. Türkiye Ormancılık Dergisi 20(3): 180-186. https://doi.org/10.18182/tjf.576898
Güney D, Atar F, Turna İ, Günlü A (2021) Effects of precommercial thinning intensity on growth of Fagus orientalis Lipsky stands over 6 years. Journal of Forestry Research https://doi.org/10.1007/s11676-021-01396-8
Güney D, Bayraktar A, Atar F, Turna İ (2020) Effects of root undercutting, fertilization and thinning on seedling growth and quality of oriental beech (Fagus orientalis Lipsky) seedlings. Artvin Çoruh Üniversitesi Orman Fakültesi Dergisi 21(2): 214-222. https://doi.org/1017474/artvinofd.696079
Güney D, Yahyaoğlu Z, Turna İ, Bayraktar A, Atar F (2019) Genetic diversity in Picea orientalis (L.) Link Populations in Different Turkey. Šumarski list 143(11-12): 539-547. https://doi.org/10.31298/sl.143.11-12.4
Güney D, Turna İ, Atar F (2018) The effects of different pretreatments on germination of Mediterranean hackberry (Celtis australis L.) seeds. Biological Diversity and Conservation 11(1): 61-67.
Güney D, Turna H, Turna İ, Kulaç Ş, Atar F, Filiz E (2016a) Variations within and among populations depending on some leaf characteristics of oriental beech (Fagus orientalis Lipsky). Biological Diversity and Conservation 9: 1-9.
Güney D, Turna İ, Bayraktar A, Seyis E, Atar E (2016b) Comparison of differently originated oriental beech (Fagus orientalis Lipsky) seedling growth in field. Biological Diversity and Conservation 9: 10-18.
Güney D, Atar F, Atar E, Turna İ, Kulaç Ş (2015) The effect of pre-treatments and seed collection time on the germination characteristics of common hornbeam (Carpinus betulus) seeds in the Eastern Black Sea Region, Turkey. Seed Science and Technology, 43(1): 1-9.
Güney D, Yahyaoglu Z, Turna İ, Müller-Strack G (2014) Genetic variation in Pinus brutia in Turkey. Fresenius Environmental Bulletion 23(5): 1249-1254.
Güney D, Bak ZD, Aydınoğlu F, Turna İ, Ayaz FA (2013) Effect of the geographical variation on the sugar composition of oriental beech (Fagus orientalis Lipsky). Turkish Journal of Agriculture and Forestry 37: 221-230. https://doi.org/10.3906/tar-1204-36
Güney D, Hatipoğlu E, Atar F, Turna İ, Kulaç Ş (2013) Changes of some morphological characteristics of oriental hornbeam (Carpinus orientalis Miller) seeds depending on altitude. International Caucasian Forestry Symposium, Artvin, Turkey, 24-26 October.
Güney D, Turna İ, Kulaç Ş (2006) Doğu kayınında (Fagus orientalis Lipsky.) vejetasyon sürelerine bağlı olarak bazı morfolojik özelliklerin irdelenmesi. Paper presented at 18. Ulusal Biyoloji Kongresi, Adnan Menderes Üniversitesi, Nobel Yayın No. 959, Fen ve Biyoloji Dizisi: 35, Kuşadası/Aydın, 26-30 Haziran.
Hamrick JL, Godt JW, Sherman-Broyles SL (1992) Factors influencing levels of genetic diversity in woody plant species. New Forests 6: 95-124.
Hamrick JL, Schnabel A (1985) Understanding the genetic structure of plant populations: Some old problems and a new approach. In: Gregorius HR (ed) Population Genetics in Forestry. Lecture Notes in Biomathematics, vol 60. Springer-Verlag, pp 50-70.
Hiraoka K, Tomaru N (2009) Population genetic structure of Fagus japonica revealed by nuclear microsatellite markers. Int J Plant Sci 170(6): 748-758.
Hiura T, Koyama H, Igarashi T (1996) Negative trend between seed size and adult leaf size throughout the geographical range of Fagus crenata. Ecoscience 3: 226-228.
Işık K (1983) Bitki gen kaynaklarımız niçin korunmalı ve planlanmalıdır? Tabiat ve İnsan 17(4): 9-15.
Işık K (1988) Orman Ağacı türlerimizde lokal ırkların önemi ve genetik kirlenme sorunları. Orman Mühendisliği Dergisi 25(11): 25-30.
Kandemir GE, Tayanç Y, Çengel B, Velioğlu E (2016) Türkiye’de yayılış gösteren kayın (Fagus) populasyonlarının moleküler filogenisi. Ormancılık Araştırma Dergisi 1(4A): 69-79. http://dx.doi.org/10.17568/oad.84722
Kaya Z (1990) Orman gen kaynaklarımız: Ulusal mirasımız. Fidan Dergisi, OGM Meslek Memurları Derneği Yayın Organı 28: 2-6.
Konnert M (1995) Investigations on the genetic variation of beech (Fagus sylvatica L.) in Bavaria. Silvae Genetica 44: 346-351.
Konnert M, Ruetz W (2003) Influence of nursery practices on the genetic structure of beech (Fagus sylvatica L.) seedling populations. Forest Ecology and Management 184(1-3): 193-200.
Koski V, Antola J (1993) Turkish national tree breeding and seed production program for Turkey (1994-2003). Coprepared with ENSO Forest Development Inc and Forest Tree Seeds and Tree Breeding Institute, Ankara.
Krzakowa M, Matras J (2005) Genetic variability among beech (Fagus sylvatica L.) populations from the Sudety Mountains, in respect of eroxidase and malate dehydrogenase loci. J Appl Genet 46(3) :271-277.
Kulaç Ş, Güney D, Turna İ (2009) Effects of seed gathering, sowing time and growing conditions on wild cherry (Prunus avium L.) seed germination. Artvin Çoruh University Faculty of Forestry Journal 10(1): 37-44.
Larsen AB (1996) Genetic structure of populations of beech (Fagus sylvatica L.) in Denmark. Scandinavian Journal of Forest Research 11(3): 220-232.
Ledig FT (1986) Conservation strategies for forest gene resources. Forest Ecology and Management 14: 77-90.
Magri D, Vendramin GG, Comps B, Dupanloup I, Geburek T, Gömöry D, Latalowa M, Litt T, Paule L, Roure JM, Tantau I, van der Knaap WO, Petit RJ, Beaulieu JL (2006) A new scenario for the Quaternary history of European beech populations: Palaeobotanical evidence and genetic consequences. New Phytol 171: 199-221.
Merzeau D, Comps B, Thıebaut B, Cuguen J, Letouzey J (1994) Genetic structure of natural stands of Fagus sylvatica L. (beech). Heredity 72: 269-277.
Mettler LE, Gregg TG (1969) Population genetics and evolution. Foundations of Modern Genetics Series, Prentice-Hall, 212 p.
Millar CI, Libby WJ (1989) Restoration: Disneyland or native ecosystem? A question of genetics. Fremontia 17(2): 3-10.
Millar CI, Marshall KA (1991) Allozyme variation of Port-Orford-Cedar (Chamaecyparis lawsoniana): Implications for genetic conservation. Forest Sci 37: 1060-1075.
Müller-Starck G (1985) Genetic differences between "tolerant" and "sensitive" beeches (Fagus sylvatica L.) in an environmentally stressed adult forest stand. Silvae Genet 34: 241-247.
Panahi P, Jamzad Z, Pourhashemi M, Hasaninejad M (2017) Morphological variation of Fagus orientalis Lipsky in the Hyrcanian forests of Iran. Iranian Journal of Botany 23(1): 37-47. https://doi.org/10.22092/ijb.2017.106702.1126
Pastorelli R, Smulders MJM, Van’t Westende WPC, Vosman B, Giannini R, Vettori C, Vendramin GG (2003) Characterization of microsatellite markers in Fagus sylvatica L. and Fagus orientalis Lipsky. Molecular Ecology Notes 3: 76-78.
Pradhan A, Yan G, Plummer JA (2004) Correlation of morphological traits with molecular markers in radish Raphanus sativus. Australian Journal of Experimental Agriculture 44: 813-819.
Rajora OP (1999) Genetic biodiversity impacts of silvicultural practices and phenotypic selection in white spruce. Theor Appl Genet 99: 954-961.
Saatçioğlu F, Ürgenç S (1960) Doğu kayını (Fagus orientalis Lipsky) tohumlarının çimlendirilmesinde soğuk ıslak işlemin etkileri üzerine araştırmalar. Orman Fakültesi Dergisi A(2): 7-26.
Sander T, König S, Rothe GM, Janben A, Weisgerber H (2000) Genetic variation of European beech (Fagus sylvatica L.) along an altitudinal transect at mount Vogelsberg in Hesse, Germany. Molecular Ecology 9: 1349-1361.
Şevik H, Ayan S, Turna İ, Yahyaoğlu Z (2010) Genetic diversity among populations in Scotch pine (Pinus sylvestris L.) seed stands of Western Black Sea Region in Turkey. African Journal of Biotechnology 9(43): 7266-7272. https://doi.org/10.5897/AJB09.1536
Shanjani PS, Vettori C, Giannini R, Khavari-Nejad RA (2004) Intraspecific variation and geographic patterns of Fagus orientalis Lipsky chloroplast DNA. Silvae Genetica 53(1-6): 193-197. https://doi.org/10.1515/sg-2004-0035
St. Clair BJ, Howe GT (2011) Strategies for conserving forest genetic resources in the face of climate change. Turkish Journal of Botany 35:403-409. https://doi.org/10.3906/bot-1012-98
Suangtho V, Graudal L, Kjaer ED (1999) Genecological zonation as a tool in conservation of genetic resources of teak (Tectona grandis) in Thailand. Journal of World Forest Resource Management 3: 15-29.
Suszka B, Kluczynska A (1980) Seedling emergence of stored beech (Fagus sylvatica L.) seed chilled without medium at controlled hydration level and pregerminated in cold-moist conditions. Arboretum Kórnickie 25: 231-255.
Takahashi M, Mukouda M, Koono K (2000) Differences in genetic structure between two Japanese beech (Fagus crenata Blume) stands. Heredity 84: 103-115.
Thomsen KA, Kjaer ED (2002) Variation between single tree progenies of Fagus sylvatica in seed traits, and its implications for effective population numbers. Silvae Genetica 51(5-6): 183-190.
Tomaru N, Takahashi M, Tsumura Y, Takahashi M, Ohba K (1998) Intraspesific variation and phylogeographic patterns of Fagus crenata (Fagaceae) mitochondrial DNA. American Journal of Botany 85(5): 629-636.
Tosun S (1993) Batı Karadeniz Bölgesi’nde doğu kayını (Fagus orientalis Lipsky) orijin denemelerinin dokuz yıllık sonuçları. Ormancılık Araştırma Enstitüsü Yayınları, Teknik Bülten No. 236-237, Ankara, pp 37-63.
Tosun S, Gülcan E (1985) Doğu kayınının (Fagus orientalis Lipsky) yapay yolla gençleştirilmesi üzerine araştırmalar. Ormancılık Araştırma Enstitüsü Yayınları, Teknik Bülten Serisi No. 133, Ankara.
Tunçtaner K (2007) Orman genetiği ve ağaç ıslahı. Türkiye Ormancılar Derneği, Eğitim Dizisi: 4, Ankara.
Turfan N, Ayan S, Nurten Yer E, Özel HB (2019) Age-related changes of some chemical components in the leaves of oriental beech (Fagus orientalis Lipsky). South-east European forestry 10(2): 117-124. https://doi.org/10.15177/seefor.19-15
Turfan N, Nurten Yer E, Ayan S (2017) The effect of magnetic field applications to chemical content of stratified seeds of oriental beech (Fagus orientalis Lipsky). Fresenius Environmental Bulletin 26(7): 4606-4615.
Tylek P, Walczyk J (2002) The relationship between the viability and geometric characteristics of beechnuts Fagus sylvatica L. El J of Polish Agr Universities, Series Forestry 5(2).
Ulaszewski B, Meger J, Mishra B, Thines M, Burczyk J (2021) Complete chloroplast genomes of Fagus sylvatica L. reveal sequence conservation in the inverted repeat and the presence of allelic variation in NUPTs. Genes 12(9): 1357.
Ürgenç S (1982) Orman ağaçları ıslahı, İ.Ü. Yayın No:2836, Orman Fakültesi Yayın No: 293, İstanbul.
Velioğlu E, Arslan ÖŞ (2000) Doğu Karadeniz göknari (Abies nordmanniana [steven] spach) ile Toros sediri (Cedrus libani a.richard) tohumlarinin tetrazolium test sonuçlarıyla çimlendirme deney sonuçlarinin mukayesesi. Orman Ağaçları ve Tohumları Islah Araştırma Müdürlüğü, Teknik Bülten No. 6, Ankara.
Velioğlu E, Çengel B, İçgen Y, Kandemir G, Kaya Z (2004) Kırklareli-Kasatura Körfezi Tabiati Koruma Alaninda bulunan karaçam (Pinus nigra Arnold) populasyonlarinin genetik yapisinin moleküler belirteçler yardimiyla belirlenmesi. Orman Ağaçları ve Tohumları Islah Araştırma Müdürlüğü, Teknik Bülten No. 15, Ankara.
Vettori C, Vendramin GG, Anzidei M, Pastorelli R, Paffetti D, Giannini R (2004) Geographic distribution of chloroplast variation in Italian populations of beech (Fagus sylvatica L.). Theor Appl Genet 109: 1-9. https://doi.org/10.1007/s00122-004-1609-9
Vornami B, Decarli N, Gailing O (2004) Spatial distribution of genetic variation in a natural beech stand (Fagus sylvatica L.) based on microsatellite markers. Conservation Genetics 5: 561-570.
Yahyaoğlu Z, Genç M (2007) Fidan standardizasyonu. Süleyman Demirel Üniversitesi, Orman Fakültesi, Yayın No. 75, Isparta.
Yalcin S (2012) Modelling the current and future ranges of Turkish pine (Pinus brutia) and oriental beech (Fagus orientalis) in Turkey in the face of climate change. Master Dissertation, Middle East Technical University, The Graduate School of Natural and Applied Sciences.
Yılmaz M (2005) Doğu kayını (Fagus orientalis Lipsky) tohumlarının fizyolojisi üzerine araştırmalar. Doctoral Dissertation, İstanbul University.
Yılmaz M (2008) Three-year storage of oriental beechnuts (Fagus orientalis Lipsky). Eur J Forest Res 127: 441-445. https://doi.org/10.1007/s10342-008-0227-5



