Schlüsselbegriffe: Populus tomentosa, Anfangsabstand; Kronenattribute; Zweigentwicklung; triploid, diploid, genetische Verbesserung, Pflanzenzüchtung
Abstract
The tree growth, crown and branch development of four Populus tomentosa clones (one diploid clone: 1316; three triploid clones: S86, B331 and B301) were examined in an 11-year-old trial plantation with seven planting density (PD) treatments ranging from 417 to 2500 stems per hectare (stems·ha-1) in northern China. Significant differences (P < 0.01) in the diameter at breast height (DBH), individual volume (V), stand volume (SV), slenderness index (SI), crown diameter (CD), ratio of CD to DBH (k/d) and live branch length (BL) were found among the PD treatments. The mean DBH, V, and CD were markedly higher under the two lowest PD treatments (417, 500 stems·ha-1) than the two highest PD treatments (1667, 2500 stems·ha-1), while SV and k/d were much higher in the two highest PD treatments. BL decreased significantly with increasing PD. Crown length (CL), the crown ratio (CR), and the numbers of all (NAB), live (NLB) and dead branches (NDB) varied in a narrow range. All growth, crown and branch development traits apart from k/d differed markedly among the four clones, demonstrating that most of these traits were under genetic control. Among the four tested clones, the three triploid clones performed better than the diploid clone for most of the traits, implying an obvious advantage of polyploidization, with B301 being the best performing triploid clone. A significant interaction was observed between the PD treatment and clone for V, SV and NAB. The findings of this study will support determining the optimal stand density and will provide scientific guidelines for the intensive management and genetic improvement of P. tomentosa plantations.
Zusammenfassung
Wachstum, Kronen- und Zweigentwicklung von vier Populus tomentosaKlonen (ein diploider Klon: 1316; drei triploide Klone: S86, B331 und B301) wurden in einer 11 Jahre alten Versuchsplantage unter sieben verschiedenen Pflanzdichten (PD) von 417 bis 2500 Stämmen pro Hektar (stems·ha-1) in Nordchina untersucht. Durchmesser in Brusthöhe (DBH), Einzelbaumvolumen (V), Bestandesvolumen (SV), Schlankheitsindex (SI), Kronendurchmesser (CD), Verhältnis von CD zu DBH (k/d) und Länge lebender Äste (BL) zeigten signifikante Unterschiede (P < 0.01) zwischen den verschiedenen PD. DBH, V und CD waren bei den beiden niedrigsten PD (417, 500 stems·ha-1) deutlich höher als bei den beiden höchsten PD (1667, 2500 stems·ha-1), wobei SV und k/d bei den beiden höchsten PD viel höher waren. Die Kronenlänge (CL), das Kronenverhältnis (CR) und die Anzahl aller Äste (NAB), die Anzahl der lebenden Äste (NLB) und die Anzahl der toten Äste (NDB), variierten hingegen wenig. Alle Merkmale des Baumwachstums, der Kronen- und Zweigentwicklung, ausgenommen k/d, zeigten bei den vier Klonen signifikante Unterschiede. Dies zeigt, dass die meisten dieser Merkmale genetisch beeinflusst sind. Von den vier untersuchten Klonen zeigten die drei triploiden Klone in den meisten getesteten Merkmalen eine bessere Leistung als der diploide Klon, was auf einen offensichtlichen Vorteil der Polyploidisierung schließen lässt. B301 war der beste triploide Klon. Es wurde eine signifikante Interaktion zwischen PD und den Klontypen V, SV und NAB beobachtet. Diese Ergebnisse erlauben eine wissenschaftlich fundierte Bestimmung angemessener Pflanzdichten und Entwicklung von Richtlinien für die intensive Bewirtschaftung von P. tomentosa-Plantagen.
1. Introduction
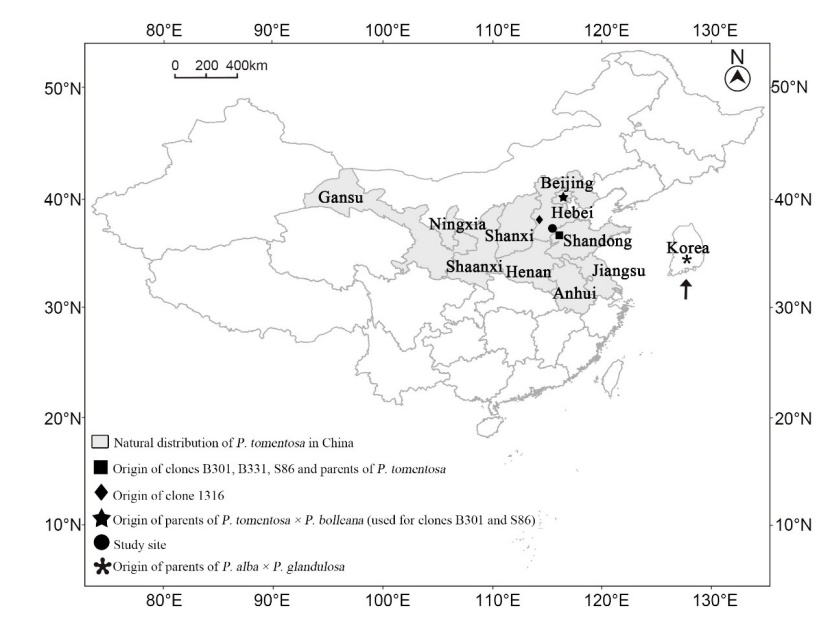
Figure 1: Natural distribution area of Populus tomentosa in China and origins of clones used in this study. / Abbildung 1: Natürliches Verbreitungsgebiet von Populus tomentosa in China und Ursprung der Klone, die in dieser Studie verwendet wurden.
Populus tomentosa is a native tree species in northern China (Du et al. 2014). It is naturally distributed along the middle and lower reaches of the Yellow River (30°N–40°N, 105°E–125°E) in northern China (Figure 1) and covers an area of approximately one million km2 (Zhang et al. 2005; Du et al. 2012). Its wood is widely used for construction, furniture and plywood production due to its straight texture, fine structure, light weight, and easy processing properties (Zhu and Zhang 1997). Populus tomentosa wood is also suitable for paper and fiber production due to its long fibers as well as high holocellulose content and low lignin content (Jin et al. 2005; Zhang et al. 2010). In the last two decades, programs aimed at germplasm selection and hybrid breeding in P. tomentosa have greatly advanced (Zhu et al. 1998; Lu et al. 2013), and considerable gains in terms of volume or biomass have been achieved through genetic improvement. However, the mean productivity of P. tomentosa plantations in China is still far lower than that of poplar plantations around the world (Xi et al. 2016). Studies on cultivation techniques are important to increase the productivity and to improve wood quality of poplar plantations (Larocque 1999). For P. tomentosa, only irrigation and fertilization have been applied so far (Xi et al. 2014; He et al. 2020), and studies on other cultivation techniques, such as planting density have not received considerable attention.
Planting density (PD) determines inter-tree competition intensity, and influences tree growth and potential timber quality (e.g. branch characteristics, knot properties and stem shape) (Medhurst et al. 2001; Gort et al. 2010). PD is the key technical parameter linked to high-yield plantations with high-quality wood production (Roth et al. 2007; Ikonen et al. 2009). The significant effects of PD on growth-related traits have been widely confirmed in a number of tree species such as P. deltoides and P. euramericana (Fang et al. 1999), Populus deltoides × P. nigra (Toillon et al. 2013), Acer velutinum (Naji et al. 2015), and Betula pendula (Niemistö 1995; Lintunen and Kaitaniemi 2010). These effects are closely related to the response of crown and branch development to PD (Hummel 2000; Pinkard and Neilsen 2003; Wang et al. 2017; 2018), since branches are the main component of the crown and supports for leaves and are responsible for nutrition and water transport among leaves as well as between leaves and stems (Ceulemans et al. 1990). The number and distribution of branches also determine the spatial distribution of leaves and, thus, crown shape and size, which affect the photosynthetic efficiency of trees (Lowell et al. 2014). Additionally, branch development directly reflects the knot properties of the stem, which determine the wood quality (Cameron and Watson 1999; Ares 2002; Mäkinen and Hein 2006; Barbour et al. 2012). As a whole, reasonable PDs play critical roles not only in making full use of forest resources and increasing stand volume and stand stability, but also in improving stem shape and reducing knot-related defects in wood.
There is abundant literature available addressing the influence of planting density on tree growth, crown and branch development, while the genetic regime has rarely been simultaneously taken into consideration, and almost none of the research in this field has involved P. tomentosa. Therefore, in the present study, growth performance and crown and branch traits were investigated in four clones of P. tomentosa, including one diploid clone and three triploid clones, under different PD treatments. The objectives were to evaluate the effects of the PD and clone on the tree growth, and crown and branch development of P. tomentosa. The findings will be contributed to the determination of reasonable planting densities, and provide evidence for the intensive plantation management of P. tomentosa and other poplar species.
2. Materials and Methods
2.1 Experimental site
The study site was located at the tree breeding base in Wei County, Hebei Province (37°2’ N, 114°18’ E), China (Figure 1). The region has a warm temperate continental semiarid monsoon climate with four distinct seasons. The mean annual temperature is 13.4°C. The mean precipitation is 584 mm and is normally concentrated from June to September. The frost-free period last on average 198 days, and the annual average sunshine duration is 2575 hours. The study site is flat with an altitude of 30-50 m.a.s.l, and the soil is a sandy loam.
2.2 Plant materials
Three triploid clones (2n = 3X = 57) and one diploid clone (2n = 2X = 38) of P. tomentosa were involved in the present study. The triploid clones were B301 ((P. tomentosa × P. bolleana) × P. tomentosa), B331 ((P. alba × P. glandulosa) × P. tomentosa) and S86 ((P. tomentosa × P. bolleana) × (P. alba × P. glandulosa)). The parents of P. tomentosa were selected in Shandong by Zhu in 1985. The parents of P. tomentosa × P. bolleana were selected in Beijing by Xu in 1958, and those of P. alba × P. glandulosa were introduced from Korea in 1984 (Wu et al. 2013). The diploid clone 1316 developed from a plus tree of P. tomentosa in Hebei Province, China (Figure 1). These clones are now widely used in commercial plantations.
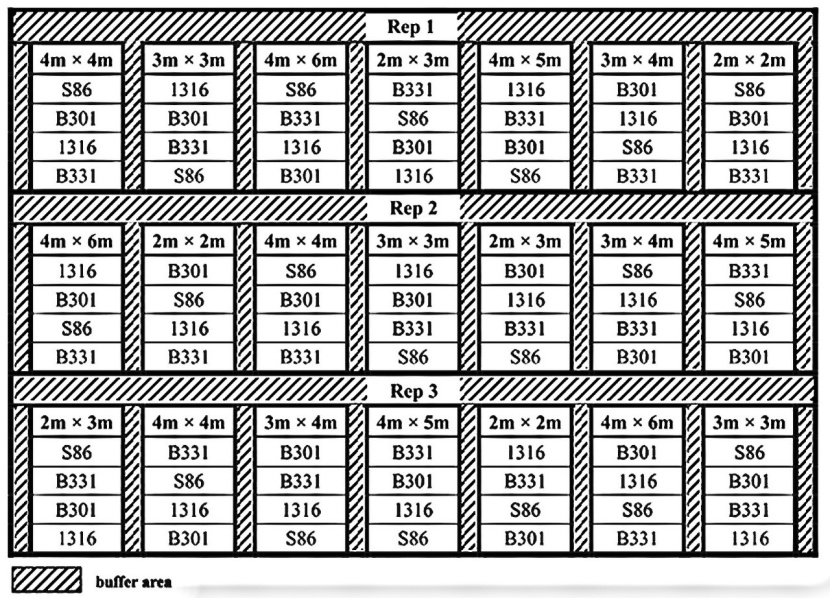
Figure 2: Plot layout for spacing trial. / Abbildung 2: Plotlayout für den Abstandsversuch.
2.3 Experimental design
The spacing trial was established in April 2007 using a split-plot, randomized complete block (RCB) design with three replicates. Seven planting density (PD) treatments (2500, 1667, 1111, 833, 625, 500 and 417 stems per hectare) were randomly allocated to the main plots and the four clones (sub-factors) were then randomly arranged to each PD treatment. In total, 84 sub-plots in 21 main plots were included in the trial (Figure 2). Each replicate was surrounded with at least two rows as a buffer area with the same clone as the nearest subplot.
2.4 Data collection
Tree height and diameter at breast height (DBH) were measured annually at the end of the growing season from 2007 until 2012 and reported earlier (Wang et al. 2012). In the present study, DBH, tree height, the height to crown base (HCB), DBH and crown diameter (CD) were measured for all trees in each plot in July 2018. Tree height, HCB and CD were measured with a Vertex IV-360 instrument (accuracy 0.01 m), and DBH was measured with diameter tape (accuracy 0.1 cm). CD was measured as the horizontal projection of the crown in four directions. A summary of all abbreviations of the measured tree attributes is given in Table 1. A mean tree was then sampled in the centre of each sub-plot with in total 84 sampled trees. The sampled trees were then felled carefully, avoiding branch loss or damage as much as possible. Their height, height to live crown base (HCB) and crown length (CL) were measured again with tape (0.1 m); live branch length (BL) and live branch diameter (BD) were measured with an electronic digital caliper (0.01 mm); and the live branch angle (BA) was measured with an electronic protractor (1°). Only the first-order branches, which were connected directly to the stem, have been considered. BD was measured as the diameter of the branch base for each live branch. This means that the largest live branch diameter (LBD) was the diameter of the largest live branch for a tree. The numbers of all (NAB), dead (NDB), and live branches (NLB) in the crown were recorded for each sampled tree. The individual stem volume (V, from tree top to ground level) was calculated on the basis of the function developed by Chen (1989) for P. tomentosa at northern China regions (Equation 1). Stand volume (SV) was the product of V multiplied by the number of stocking per hectare (N) (Equation 2). The slenderness index (SI) was the ratio of height to DBH (Equation 3). The ratio of CD and DBH (k/d) and the crown ratio (CR) were calculated using Equations 4 and 5 below, respectively.

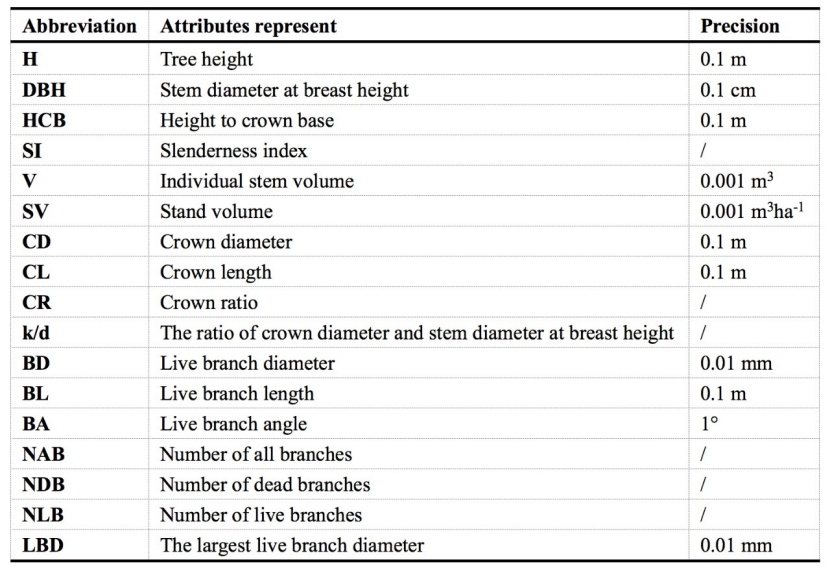
Table 1: Explanation of symbols and abbreviations. / Tabelle 1: Erläuterung der Symbole und Abkürzungen.
2.5 Statistical analysis of the data
The growth (i.e., H, DBH, V, SV, HCB, SI) and crown (i.e., CD, CL, CR, k/d) traits as well as survival rate were analyzed using analysis of variance (ANOVA) for a split-plot design and Tukey’s multiple range tests. Prior to the analyses, the data of survival rate was arcsine transformed. The differences in the branch number (i.e., NAB, NLB, NDB) and morphology traits (i.e., BL, BD, BA) were analyzed using linear mixed models (6) and (7) below, with PD and clone as fixed effects and the block or the block and tree as random effects, respectively.

where ydcb and ydcbt, the observed value; μ , the overall mean; βd, effect of PD; βc, effect of clone; μb, random effect for block; μbt, random effect for tree; and ε, the residual error. All statistical analyses were conducted with R statistical software (R 3.5.2).
3. Results
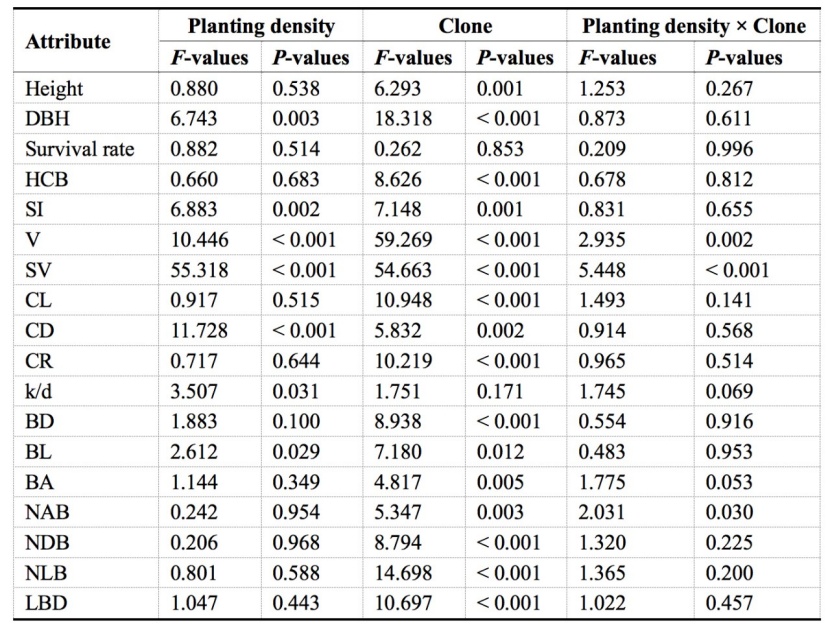
Table 2: Effects of planting density, clone and their interaction (planting density × clone) on tree growth, crown and branch properties Populus tomentosa / Tabelle 2: Auswirkungen der Pflanzdichte, des Klontyps und deren Interaktion (Pflanzdichte × Klon) auf Baumwachstum, Kronen- und Zweigeigenschaften bei Populus tomentosa.
The results indicated that diameter at breast height (DBH), individual volume (V), stand volume (SV), the slenderness index (SI), crown diameter (CD), the ratio of CD to DBH (k/d), and live branch length (BL) differed significantly (P < 0.01) among the planting density (PD) treatments. Almost all traits differed markedly among the four clones except for k/d and the survival rate; and only V, SV and the number of all branches (NAB) showed a significant interaction between the PD treatment and clone (Table 2).
3.1 Tree growth performance
Mean DBH and V of 11-year-old P. tomentosa decreased, while SI and SV increased with increasing PD (Table 3). DBH of the 2500 stems·ha-1 treatment was significantly lower than all other PDs, while DBH of the 1667 stems·ha-1 treatment was still significantly lower than DBH of the two lowest PD (500 and 417 stems·ha-1). DBH reached a peak in the 500 stems·ha-1 treatment, but it did not differ significantly from those of the 833 and 417 stems·ha-1 treatments. V showed similar differences to DBH among the PD treatments, while SV and SI were the highest in the 2500 stems·ha-1 treatment, and was significantly higher than those under the four lower PD treatments. H, HCB and survival rate ranged from 20.0 to 21.8 m, from 6.4 to 8.0 m and from 85 % to 92 %, respectively, in the four P. tomentosa clones at the age of 11 years, and significant differences were absent among the PD treatments (P ≥ 0.05) (Table 3).
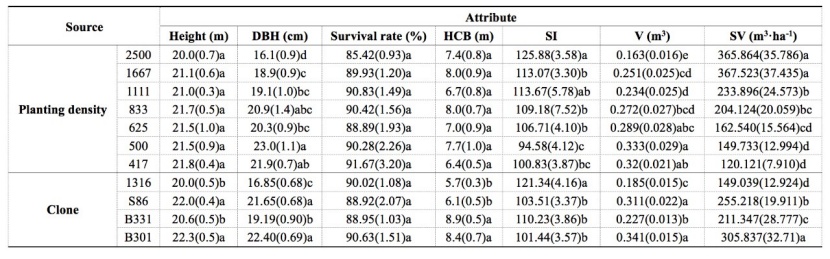
Table 3: Growth performance of 11-year-old Populus tomentosa plantations in response to planting density. Numbers in the parentheses are standard error of mean value. Means within the same row marked with the same lowercase letters are not significantly different (P ≥ 0.05). / Tabelle 3: Wachstumsleistung von 11-jährigen Populus tomentosa Plantagen in Abhängigkeit von der Pflanzdichte. Die Zahlen in Klammern sind Standardfehler des Mittelwerts. Mittelwerte innerhalb derselben Zeile, die mit denselben Kleinbuchstaben markiert sind, unterscheiden sich nicht signifikant (P ≥ 0.05).
Growth performance differed greatly among the clones. H, DBH, V, SV and HCB of diploid clone 1316 were generally much lower, while the SI of this clone was significantly higher than those of triploid clones S86, B331 and B301. Among the three triploid clones, B301 performed the best in terms of H, DBH, V and SV, followed by S86 (Table 3).
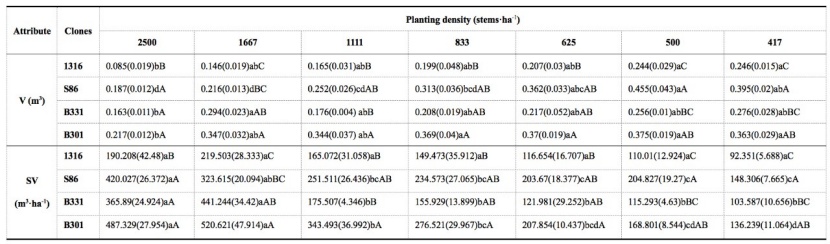
Table 4: Comparison of individual volume (V) and stand volume (SV) for four clones of Populus tomentosa under different planting density treatments. We show means and standard errors in brackets; lowercase letters represent significant difference between planting densities for the same clone, and capital letters represent the difference between clones under the same planting density treatment. Absent significant difference between paired treatments or clones are indicated by the same letters (P ≥ 0.05). / Tabelle 4: Vergleich des Einzelvolumen (V) und des Ständervolumen (SV) für vier Klone von Populus tomentosa unter verschiedenen Pflanzdichten. Daten werden als Mittelwerte angezeigt (Standardfehler in Klammer); Kleinbuchstaben zeigen signifikante Unterschiede zwischen Pflanzdichten für denselben Klon und Großbuchstaben stellen den Unterschied zwischen Klonen unter derselben Pflanzdichtebehandlung dar. Es liegt kein signifikanter Unterschied zwischen gepaarten Behandlungen oder Klonen bei denselben Buchstaben vor (P ≥ 0.05).
Since V and SV were significantly affected by the interaction of PD and clone, the differences among clones in each PD treatment were further analyzed. The V of all four clones declined, and SV increased significantly as PD increased. For any given PD treatment, the V and SV of diploid clone 1316 were markedly lower than those of triploid clones B331, B301 and S86, and a significant difference was always found between diploid clone 1316 and triploid clone B301. For the other two triploid clones, the V and SV of S86 were much higher than those of B331 under all PD treatments except 1667 stems·ha-1. The highest V (0.455 m3) was observed under the 500 stems·ha-1 treatment in triploid clone S86, and was 4.35 times greater than that of diploid clone 1316 under the 2500 stems·ha-1 treatment. The highest SV (520.621 m3·ha-1) was found under the 2500 stems·ha-1 treatment in triploid clone B301, which was 4.4 times greater than that of diploid clone 1316 under the 417 stems·ha-1 (Table 4).
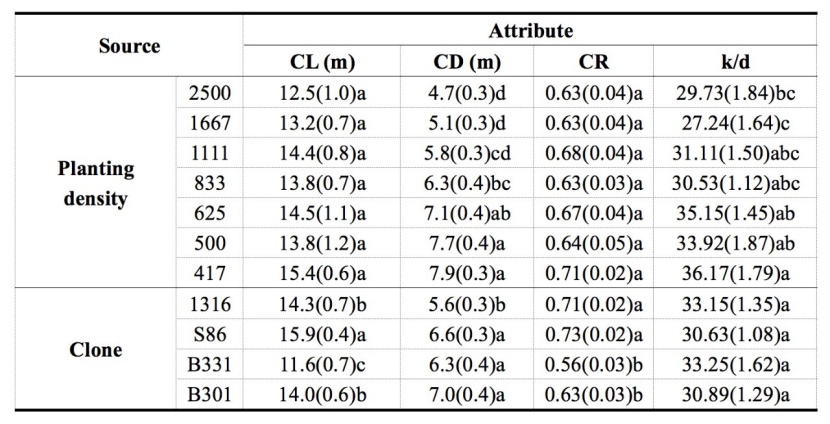
Table 5: The crown attributes of 11-year-old Populus tomentosa plantations under seven planting density treatments. For details please see Table 3. / Tabelle 5: Die Kroneneigenschaften bei 11-jährigen Populus tomentosa-Plantagen mit sieben Pflanzdichtebehandlungen. Für Details verweisen wir auf Tabelle 3.
3.2 Crown attributes
The crown length (CL), crown diameter (CD), crown ratio (CR) and k/d were presented in Table 5 for the 11-year-old P. tomentosa clones under different PD treatments. CL increased slightly from 12.5 m in the 2500 stems·ha-1 treatment to 15.4 m in the 417 stems·ha-1 treatment. CR showed slight fluctuation around 0.66. CD increased markedly with decreasing PD and differed significantly between the high-PD (2500, 1667 stems·ha-1) and low-PD (625, 500, 417 stems·ha-1) treatments. The CD of the lowest PD treatment was approximately 68.1 % higher than that of the highest PD treatment. There was an obvious increasing trend of k/d with decreasing PD, and significant differences were observed between the lowest (417 stems·ha-1) and the two highest (1667 and 2500 stems·ha-1) PD treatments.
Regarding the variation in crown traits among the four P. tomentosa clones (Table 5), diploid clone 1316 showed no significant difference in CL from triploid clone B301, while their CLs were significantly higher than that of triploid B331, but lower than that of triploid clone S86. The CRs of clones 1316 and S86 were markedly higher than those of clones B331 and B301. The CDs of the three triploid clones were significantly higher than that of diploid clone 1316. Significant difference was absent in k/d among the four clones.
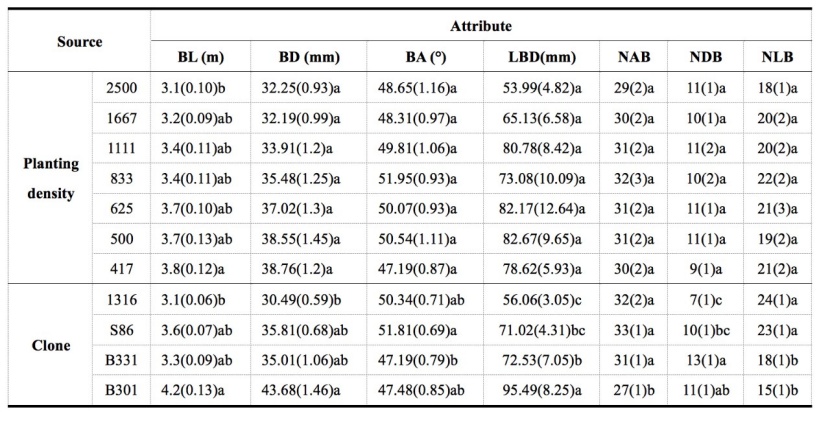
Table 6: The branch attributes of 11-year-old Populus tomentosa plantations under seven planting density treatments. For details please see table 3. / Tabelle 6: Die Zweigmerkmale bei 11-jährigen Populus tomentosa-Plantagen mit sieben Pflanzdichtebehandlungen. Für Details verweisen wir auf Tabelle 3.
3.3 Branch Attributes
Among the seven tested traits related to branch development, only the live branch length (BL) differed significantly among the seven PD treatments, and a considerable difference was observed between the highest (2500 stems·ha-1) and lowest (417 stems·ha-1) PD treatments in the 11-year-old trees (Table 6). However, increasing trends with decreasing PD were observed for the mean live branch diameter and the largest live branch diameter although significant difference was absent in them among PD treatments.
Table 6 also showed that branch development differed significantly among the four P. tomentosa clones. The BL, live branch diameter (BD), and the largest live branch diameter (LBD) of the triploid clones were larger than those of diploid clone 1316, with B301 showing the highest values among all four clones. For the live branch angle (BA), significant difference was absent between the triploid clones and diploid clone, but a significant difference was seen between triploid clones B331 and S86.
The numbers of all branches (NAB) and dead branches (NDB) in the crown were approximately 30 and 10, respectively, regardless of the PD treatment (Table 6). However, significant differences in NAB, number of live branches (NLB), and NDB were found among the four clones. The NAB of clone B301 was markedly lower than those of clones S86, B331, and 1316. The NDBs of clones B331 and B301 were significantly higher than that of clone 1316. The NLBs of clones 1316 and S86 were significantly higher than those of clones B331 and B301.
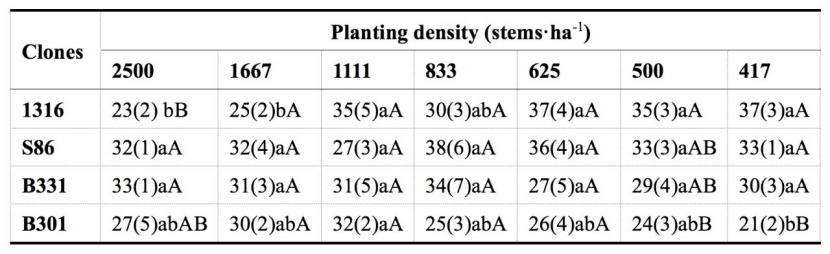
Table 7: Comparison on number of all branches for four clones of Populus tomentosa under different planting density treatments. For details please see Table 4. / Tabelle 7: Vergleich der Anzahl aller Zweige für vier Klone von Populus tomentosa mit verschiedenen Pflanzdichtebehandlungen. Für Details verweisen wir auf Tabelle 4.
The NABs of the different clones responded differently to the PD treatments. Significant differences among planting densities were only observed for diploid clone 1316 and triploid clone B301. NAB under the lower PD treatments (417, 500 and 625 stems·ha-1) was considerably higher than that under the higher PD treatments (2500 and 1667 stems·ha-1) for clone 1316, while for clone B301, the 1111 stems·ha-1 treatment showed the highest NAB, which was significantly higher than that under the lowest PD treatment (417 stems·ha-1). Regarding the NAB differences among clones, only the highest PD treatment and the two lowest PD treatments showed notable differences. Under the 2500 stems·ha-1 treatment, the NAB of clones S86 and B331 was considerably greater than that of clone 1316, while the NAB of clone 1316 was significantly higher than that of clone B301 under the 500 and 417 stems·ha-1 treatments (Table 7).
4. Discussion
4.1 Effect of planting density
In this study, diameter at breast height (DBH) and individual volume (V) were negatively influenced by the planting density (PD), while slenderness index (SI) and stand volume (SV) were significantly positively influenced. Tree height on the other hand was not markedly affected by PD. These findings are consistent with previous studies on Populus simonii × P. nigra (Jiang et al. 2007), Eucalyptus pilularis (Alcorn et al. 2007), Fraxinus excelsior and Acer pseudoplatanus (Hein and Spiecker 2008), but contradict studies on Populus trichocarpa (Heilman and Peabody 1981) and Populus alba var. pyramidalis (Huang et al. 2017), where DBH and height were all markedly influenced by PD. The disagreement with these studies indicated that the effects of PD on height growth are more complicated and height growth is affected by multiple factors, such as species, age, site conditions, and the range of PD treatments (Daniel et al. 1979; Sun 1992; Hummel 2000). In an earlier study of the trial examined here, the height of P. tomentosa was significantly influenced by PD at the age of 2-3.5 years, while this effect was absent when the trees were four years old (Wang et al. 2012). It could thus be deduced that P. tomentosa showed notable differences in height growth only before 4 years of age, when canopy closure occurred. P. tomentosa is characterized by fast growth; for example, the rapid height growth stage of its triploid occurs before 8 years of age, and its height growth rate decreases thereafter (Zhu 2006), when less of a difference may be observed between PD treatments. This could also explain why significant difference was absent in height among the seven PD treatments at the age of 11-year-old trees in the present study.
Similar to DBH, crown diameter (CD) and the ratio of CD to DBH (k/d) were significantly negatively influenced by PD, while crown length (CL) and the crown ratio (CR) were not in the present study. These results differed to some extent from Akers et al.’s (2012) study on 12-year-old Pinus taeda plantations, which showed that CD and CL decreased markedly with increasing PD. These results were also slightly inconsistent with Wang et al.’s (2017) study on Betula alnoides, they found that CD, CL and CR differed considerably between the lowest PD treatment and the other four treatments, while significant difference was absent in k/d. In the present study, the CDs under the higher PD treatments (1111, 1667 and 2500 stems per hectare (stems·ha-1)) were much lower than those under the lower PD treatments, and only the CD under the lowest PD treatment (417 stems·ha-1) differed markedly from those under the two highest PD treatments (1667 and 2500 stems·ha-1). These discordances could be explained by the differences in the species involved in these studies. B. alnoides is a valuable species with a rotation period of more than 20 years, while P. tomentosa is a fast-growing species with a short rotation period of 5 to 10 years. These discordances might also result from differences in age. Johnson et al. (2015) found that the CD and CL of Pinus taeda differed significantly among different PD treatments at the age of 14 years but not at the age of 15 years.
Wang et al.’s (2017) study on B. alnoides showed that the live branch diameter (BD) and largest live branch diameter (LBD) of 14-year-old B. alnoides tended to decrease significantly with increasing PD, and the numbers of all (NAB), live (NLB) and dead branches (NDB) varied within narrow ranges. In the present study, only the live branch length (BL) of P. tomentosa was significantly affected by PD. The discordance of both studies might be caused by the fact that inter-tree competition for resources such as light and space intensified as tree age increased, which resulted in decreases in BL, BD and LBD with increasing PD. It was inferred that the BL of P. tomentosa showed quite strong plasticity in response to inter-tree competition compared to BD.
4.2 Effect of clone
Almost all examined traits differed significantly among the clones in the present study except for k/d, which was influenced greatly by PD rather than the clone. These results were in accordance with those of most previous studies that the branch attributes of P. tomentosa were mainly controlled by genetic factors in the present study. Taking branch traits as an example, BL, NAB and the live branch angle (BA) were also differed markedly among the poplar clones (Nelson et al. 1981; Ceulemans et al. 1990; Benomar et al. 2012). Some studies have also shown that NAB (Mäkinen et al. 2001) and BA (Vestøl et al. 1999) are subject to genetic control at moderate or strong levels. Among the four clones included in the present study, the three triploids performed better than the diploid in terms of height, DBH, HCB, SI, V, SV CD, BL, BD and LBD, demonstrating an obvious advantage of polyploidization. Diploid clone 1316 exhibited the most promising branch diameters values, but the lowest volume growth. The triploid clones exhibited higher volume production than the diploid clone. Given that the triploid clones performed better in most traits, they should be applied preferentially. Considering their branch diameter is larger, artificial pruning can be used to decrease related defects. B301 was the best triploid clone in terms of growth performance; however, given the effect of branch traits on knot-related defects, it is necessary to control the branches within the target height by artificial pruning in the management of plantations to meet multiple utilization requirements for P. tomentosa. Additionally, the considerable variation in most growth, crown and branch traits among the four P. tomentosa clones in the present study partly explains the insignificant differences among PD treatments.
5. Conclusion
Planting density (PD) was negatively influenced tree growth but did not significantly affect branch development; triploid clones showed great advantages in growth performance and exhibited larger branches than diploid clones, and there were significant interactions between the PD and clone in terms of individual volume, stand volume and the number of all branches. Therefore, the determination of optimal planting densities for different clones is critical for the efficient cultivation of P. tomentosa, based on the final wood production target. The pulpwood cultivation of P. tomentosa is suggested to be conducted under a higher planting density, such as 1667 stems·ha-1, especially for triploid clone B301 (showing the highest SV). For high-quality sawn timber production, planting the P. tomentosa triploid clone S86 (showing the highest V and SV, smaller BD) under a lower density (e.g., 500 stems·ha-1) would be much suitable.
Acknowledgments
We specifically express our deep remembrance to Professor Dr. Zhiyi Zhang. He designed the planting density experiment and established the trial plantations. This study was supported by the National Key Research and Development Program of China (2016YFD0600403).
Conflict of interest
The authors declare no conflict of interest.
Reference
Akers M., Kane M., Teskey R., Daniels R., Zhao D., Subedi S. 2012. The effects of planting density and cultural intensity on loblolly pine crown characteristics at age twelve. In: Butnor JR (eds) Proceedings of the 16th biennial southern silvicultural research conference. e-Gen. Tech. Rep. SRS-156. Asheville, NC: US Department of Agriculture Forest Service, Southern Research Station, pp 1-1.
Alcorn PJ., Pyttel P., Bauhus J., Smith RGB., Thomas D., James R., Nicotra A. 2007. Effects of initial planting density on branch development in 4-year-old plantation grown Eucalyptus pilularis and Eucalyptus cloeziana trees. Forest Ecology and Management, 252(1-3): 41-51.
Ares A. 2002. Changes through time in traits of poplar clones in selection trials. New Forests, 23(2): 105-119.
Barbour RJ., Kellogg RM. 1990. Forest management and end-product quality: a Canadian perspective. Canadian Journal of Forest Research, 20(4): 405-414.
Benomar L., DesRochers A., Larocque GR. 2012. The effects of spacing on growth, morphology and biomass production and allocation in two hybrid poplar clones growing in the boreal region of Canada. Trees, 26(3): 939-949.
Cameron AD., Watson BA. 1999. Effect of nursing mixtures on stem form, crown size, branching habit and wood properties of Sitka spruce (Picea sitchensis (Bong.) Carr.). Forest Ecology and Management, 122(1-2): 113-124.
Ceulemans R., Stettler RF., Hinckley TM., Isebrands JG., Heilman PE. 1990. Crown architecture of Populus clones as determined by branch orientation and branch characteristics. Tree Physiology, 7(1-2-3-4): 157-167.
Chen ZS. 1989. The establishment of binomial stock volume tables of poplar. Forest Research, 2(1): 78-83. (In Chinese with English Abstract)
Daniel TW., Helms JA., Baker FS. 1979. Principles of silviculture Mc-Graw-Hill Book Co., USA.
Du QZ., Wang BW., Wei ZZ., Zhang DQ., Li BL. 2012. Genetic diversity and population structure of Chinese white poplar (Populus tomentosa) revealed by SSR markers. Journal of Heredity, 103(6), 853-862.
Du QZ., Xu BH., Gong CR., Yang XH., Pan W., Tian JX., Li BL., Zhang DQ. 2014. Variation in growth, leaf, and wood property traits of Chinese white poplar (Populus tomentosa), a major industrial tree species in Northern China. Canadian Journal of Forest Research, 44(4): 326-339.
Fang SZ., Xu XZ., Lu SX., Tang LZ. 1999. Growth dynamics and biomass production in short-rotation poplar plantations: 6-year results for three clones at four spacings. Biomass Bioenergy, 17(5): 415-425.
Gort J., Zubizarreta-Gerendiain A., Peltola H., Kilpeläinen A., Pulkkinen P., Jaatinen R., Kellomäki S. 2010. Differences in branch characteristics of Scots pine (Pinus sylvestris L.) genetic entries grown at different spacing. Annals of Forest Science, 67(7): 705.
He YL., Xi BY., Bloomberg, M., Jia, LM., Zhao, DH. 2020. Effects of drip irrigation and nitrogen fertigation on stand growth and biomass allocation in young triploid Populus tomentosa plantations. Forest Ecology and Management, 461, 117937.
Heilman P., Peabody Jr DV. 1981. Effect of harvest cycle and spacing on productivity of black cottonwood in intensive culture. Canadian Journal of Forest Research, 11(1): 118-123.
Hein S., Spiecker H. 2008. Crown and tree allometry of open-grown ash (Fraxinus excelsior L.) and sycamore (Acer pseudoplatanus L.). Agroforestry systems, 73(3): 205-218.
Huang CX., Zhang M., Ba YH., Zhang HJ., Kang YX., Wang YC. 2017. Effect of planting density on the growth and photosynthetic characteristics of Xinjiang poplar plantation. Journal of Gansu Agricultural University, 52(2): 61-64. (In Chinese with English Abstract)
Hummel S. 2000. Height, diameter and crown dimensions of Cordia alliodora associated with tree density. Forest Ecology and Management, 127(1-3): 31-40.
Ikonen V., Kellomäki S., Peltola H. 2009. Sawn timber properties of Scots pine as affected by initial stand density, thinning and pruning: a simulation based approach. Silva Fennica, 43: 411-431.
Jiang ZH., Wang XQ., Fei BH., Ren HQ., Liu XE. 2007. Effect of stand and tree attributes on growth and wood quality characteristics from a spacing trial with Populus xiaohei. Annals of forest science, 64(8): 807-814.
Jin XJ., Pu JW., Xie YM., Takeshi F., Liu XY. 2005. Lignin characterization of triploid clones of Populus tomentosa Carr. Forestry Studies in China, 7(2): 52-56.
Johnson E., Kane M., Zhao D. 2015. Cultural intensity and planting density effects on individual tree-stem growth, stand and crown attributes, and stand dynamics of non-thinned and thinned loblolly pine plantations during the age 12-to 15-year period in the upper coastal plain and piedmont of the southeastern United States. In: Holley AG, Connor KF., Haywood JD (eds) Proceedings of the 17 th biennial southern silvicultural research conference. e-Gen. Tech. Rep. SRS-203. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station, pp 362-363.
Larocque GR. 1999. Performance and morphological response of the hybrid poplar DN-74 (Populus deltoides × nigra) under different spacings on a 4-year rotation. Annals of Forest Science, 56(4), 275-287.Lintunen A., Kaitaniemi P. 2010. Responses of crown architecture in Betula pendula to competition are dependent on the species of neighbouring trees. Trees, 24(3): 411-424.
Lowell E., Maguire D., Briggs D., Turnblom E., Jayawickrama K., Bryce J. 2014. Effects of silviculture and genetics on branch/knot attributes of coastal pacific northwest douglas-fir and implications for wood quality-A Synthesis. Forests, 5(7): 1717-1736.
Lu M., Zhang PD., Kang XY. 2013. Induction of 2n female gametes in Populus adenopoda Maxim by high temperature exposure during female gametophyte development. Breeding science, 63(1): 96-103.
Mäkinen H., Hein S. 2006. Effect of wide spacing on increment and branch properties of young Norway spruce. European Journal of Forest Research, 125(3): 239-248.
Mäkinen H., Saranpää P., Linder S. 2001. Effect of nutrient optimization on branch characteristics in Picea abies (L.) Karst. Scandinavian Journal of Forest Research, 16(4): 354-362.
Medhurst JL., Beadle CL., Neilsen WA. 2001. Early-age and later-age thinning affects growth, dominance, and intraspecific competition in Eucalyptus nitens plantations. Canadian Journal of Forest Research, 31(2): 187-197.
Naji HR., Nia MF., Kiaei M., Abdul-Hamid H., Soltani M., Faghihi A. 2015. Effect of intensive planting density on tree growth, wood density and fiber properties of maple (Acer velutinum Boiss.). iForest-Biogeosciences Forestry, 9(2): 325.
Nelson ND., Burk T., Isebrands JG. 1981. Crown architecture of short-rotation, intensively cultured Populus.: I. Effects of clone and spacing on first-order branch characteristics. Canadian Journal of Forest Research, 11(1): 73-81.
Niemistö P. 1995. Influence of initial spacing and row‐to‐row distance on the growth and yield of silver birch (Betula pendula). Scandinavian Journal of Forest Research, 10(1-4): 245-255.
Pinkard EA., Neilsen WA. 2003. Crown and stand characteristics of Eucalyptus nitens in response to initial spacing: implications for thinning. Forest Ecology and Management, 172(2-3): 215-227.
Roth BE., Li X., Huber DA., Peter GF. 2007. Effects of management intensity, genetics and planting density on wood stiffness in a plantation of juvenile loblolly pine in the southeastern USA. Forest Ecology and Management, 246(2-3): 155-162.
Sun SX. 1992. Silviculture. China Forestry Publishing House, Beijing.
Toillon J., Fichot R., Dallé E., Berthelot A., Brignolas F., Marron N. 2013. Planting density affects growth and water-use efficiency depending on site in Populus deltoides × P. nigra. Forest Ecology and Management, 304: 345-354.
Vestøl GI., Colin F., Loubère M. 1999. Influence of progeny and initial stand density on the relationship between diameter at breast height and knot diameter of Picea abies. Scandinavian Journal of Forest Research, 14(5): 470-480.
Wang CS., Tang C., Hein S., Guo JJ., Zhao ZG., Zeng J. 2018. Branch development of five-year-old Betula alnoides plantations in response to planting density. Forests, 9(1): 42.
Wang CS., Zeng J., Hein S., Zhao ZG., Guo JJ., Zeng J. 2017. Crown and branch attributes of mid-aged Betula alnoides plantations in response to planting density. Scandinavian Journal of Forest Research, 32(8): 679-687.
Wang LB., Zhang ZY., Kang XY., Shen YB., Song LJ., Zhang YH. 2012. The effect of planting density on heritability of growth traits in Populus tomentosa Clones. Chinese Agricultural Science Bulletin, 28(10): 13-20. (In Chinese with English Abstract)
Wu F., Zhang PD., Pei JC., Kang XY. 2013. Genotypic parameters of wood density and fiber traits in triploid hybrid clones of Populus tomentosa at five clonal trials. Annals of forest science, 70(7), 751-759.
Xi BY., Li GD., Bloomberg M., Jia LM. 2014. The effects of subsurface irrigation at different soil water potential thresholds on the growth and transpiration of Populus tomentosa in the North China Plain. Australian forestry, 77(3-4), 159-167.
Xi BY., Bloomberg M., Watt MS., Wang Y., Jia LM. 2016. Modeling growth response to soil water availability simulated by HYDRUS for a mature triploid Populus tomentosa plantation located on the North China Plain. Agricultural Water Management, 176, 243-254.
Zhang Q., Zhang ZY., Lin SZ., Lin YZ. 2005. Resistance of transgenic hybrid triploids in Populus tomentosa Carr. against 3 species of lepidopterans following two winter dormancies conferred by high level expression of cowpea trypsin inhibitor gene. Silvae genética, 54(1-6), 108-116.
Zhang AP., Liu CF., Sun RC. 2010. Fractional isolation and characterization of lignin and hemicelluloses from Triploid of Populus tomentosa Carr. Industrial crops and products, 31(2): 357-362.
Zhu ZT., Kang XY., Zhang ZY. 1998. Studies on selection of natural triploids of Populus tomentosa. Scientia Silvae Sinicae, 34(4): 22-31. (In Chinese with English abstract)
Zhu ZT., Zhang ZY. 1997. The status and advances of genetic improvement of Populus tomentosa Carr. Journal of Beijing Forestry University, 6(1): 1-7.
Zhu ZT. 2006. Genetic improvement of Populus tomentosa Carr., China Forestry Publishing House, Beijing.



