Schlüsselbegriffe: Larix gmelinii, Betula platyphylla, Permafrost, Waldbrand, Holznutzung, Bodentemperatur, Bodenfeuchte, Kohlenstoffsequestrierung
Abstract
Management of post-fire forest is essential for carbon cycling and forest ecosystem. Effects of post-fire logging on soil respiration have not been well studied, in particular in permafrost regions. We examined soil respiration rate, soil temperature and soil moisture during two growing seasons within the permafrost area of Daxing’anling in northeastern China. We were interested in the effects on soil respiration of (1) the forest fire and (2) the post-fire salvage logging. Soil respiration was measured by applying LI-8100 (LI-COR Inc.) measurement system. Heterotrophic soil respiration was measurement with the trenching method that cut off the roots resulting in root death without removing them. Our results indicated that the autotrophic soil respiration significantly increased after fire (P < 0.05). Selective-cutting increased both total soil respiration and heterotrophic soil respiration, while clear-cutting reduced soil respiration and both respiration components. Soil respiration of the selective-cut plot were significantly higher than that of the clear-cut plot (P < 0.05). Fire was associated with an increase in the share of autotrophic soil respiration, while salvage-logging of post-fire forest reduced this share, as the ratio of autotrophic soil respiration in the clear-cut plot was significantly lower than that in the unlogged area (P < 0.05). With increasing salvage logging intensity after fire disturbance, soil temperature increased. The varying and somewhat contradicting effects of different logging practices on soil respiration in burnt forest ecosystems require more studies to disentangle the effect of salvage logging and fire on soil respiration.
Zusammenfassung
Die Bewirtschaftung nach einem Waldrand ist wichtig für den Kohlenstoffkreislauf von Waldökosystemen. Die Auswirkungen von Holznutzung nach einem Feuer auf die Bodenatmung sind insbesonders für Permafrostregionen nicht gut untersucht. Wir haben die Bodenatmung, Bodentemperatur und Bodenfeuchtigkeit während zwei Vegetationsperioden im Permafrostgebiet von Daxing'anling im Nordosten Chinas gemessen, um die Auswirkungen von Waldbränden und Holznutzung nach einem Brand auf die Bodenatmung zu verstehen. Die Bodenatmung wurde mit einem LI-8100 (LI-COR Inc.) gemessen. Die heterotrophe Bodenatmung wurde mit der Grabenmethode gemessen, bei der durch Abtrennen die Wurzeln zum Absterben gebracht, aber nicht entfernt werden. Unsere Ergebnisse zeigten, dass die autotrophe Bodenatmung nach dem Waldbrand signifikant zunahm (P < 0.05). Die selektive Nutzung von verbrannten Bäumen erhöhte sowohl die gesamte Bodenatmung als auch die heterotrophe Bodenatmung, während Kahlschlag die Bodenatmung generell reduzierte. Die Bodenatmung auf selektiv genutzten Flächen war deutlich höher als jene von Kahlschlagflächen (P < 0.05). Waldbrand führte zu einem Anstieg des Anteils der autotrophen Bodenatmung, während die Nutzung nach dem Waldbrand die autotrophe Bodenatmung reduzierte und der Anteil der autotrophen Bodenatmung bei Kahlschlag deutlich niedriger als in nicht genutzten Beständen war (P < 0.05). Der Waldbrand führte zu einer Erhöhung der Bodentemperatur, die mit Intensität der Nutzung weiter anstieg. Die unterschiedlichen und teils widersprüchlichen Auswirkungen verschiedener Nutzungsformen auf die Bodenatmung in verbrannten Wälder erfordern weitere Studien zur Entflechtung der Auswirkungen von Waldbrand und Nutzung auf die Bodenatmung.
1. Introduction
Forests are the largest terrestrial ecosystem and slow down the pace of global warming by regulating global carbon balance and controlling the increase of atmospheric concentrations of greenhouse gases. Wildfire is a globally important disturbance in forest ecosystems. Many studies have shown that fire plays a crucial role in forest carbon fluxes (e.g. Kelly et al., 2016; Koster et al., 2018). The increasing surface temperature after fire disturbance has been reported to accelerate thawing of frozen soil in boreal forests, increase soil respiration, and ultimately lead to soil carbon loss (Aaltonen et al., 2019). After a forest fire, burnt trees remain dead standing in the forest and by occupying large areas continue to affect the soil carbon cycle. Accordingly, various post-fire management techniques (e.g., clear-cutting, selective-logging, afforestation) were used to restore the burnt forests, which inevitably affects soil respiration (Parro et al., 2019; Wittenberg et al., 2020). To recover economic losses and finance revegetation of the burned area, often post-fire managements involve ‘salvage logging of the burned forest’ (Garcia-Carmona et al., 2020; Martínez-García et al., 2017). However, such interventions in burnt forests have not been justified taking into account the effects on carbon dynamics and/or soil respiration.
Concilio et al. (2005) found that the soil respiration of mixed coniferous and broadleaved forests increased after logging and burning, but the effect of burning was not significant. Salvage logging is a secondary disturbance to a post-fire forest, resulting in further reductions of tree biomass, a decrease in forest canopy density and an increase in surface radiation, which should affect the soil carbon cycle (Serrano-Ortiz et al., 2011; Stevens and Van, 2008). Lopez-Serrano et al. (2016) found that soil respiration increased after thinning on fire in mixed Mediterranean forests. Parro et al. (2019) reported soil respiration in burned and cleared areas was not reduced compared to burned and uncleared areas, while the soil respiration in unburned control areas was two times higher than that in burned areas. Therefore, both burning wood and cutting wood may have an effect on soil respiration in forest areas. Both forest fire and post-fire logging management could affect soil temperature, humidity, and nutrient content, which led to soil respiration volatility in forests and woodland (Hu et al., 2016; Lopez–Serrano et al., 2016).
The variability in soil respiration had been reported to be caused by changing soil temperature and soil respiration would raise exponentially with increasing soil temperature, given sufficient soil moisture (Hubbard et al., 2005). Eom et al. (2018) found that when the air temperature was high, the effect of precipitation on soil respiration was higher than that of soil temperature. Therefore, a regression model of soil respiration may provide a prediction of soil respiration dynamics, when considering the dominant factors such as temperature and moisture (Song et al., 2018). The combustion of organic matter and tree mortality caused by fire can lead to a decline in tree growth and reduction of soil autotrophic respiration (Tang et al., 2020). The release of soil CO2 in forests after logging is affected by many factors, including regional climatic condition, local weather condition, soil carbon stock, groundwater level, tree age, species composition, local topography and logging intensity (Molchanov et al., 2017).
Clear-cutting may have a higher degree of interference with the accumulation of organic matter in mature trees than the fire disturbance, while the fire has greater effects on vegetation composition and soil structure (Haeussler and Bergeron, 2004). However, there are still controversies about the potential negative effects of post-fire logging management, regarding its impacts on vegetation restoration, soil properties and soil microbial characteristics (Francos et al., 2018). Lafleur et al. (2016) studied the effects of logging and fire disturbance on the northern forest and pointed out that fire is a ‘chemical disturbance’ while logging is a ‘mechanical disturbance’. In the logging process, the forest floor is mechanically compacted. Furthermore, nutrients released by forest fires can be absorbed by the regenerating vegetation (Thiffault et al., 2007). The effects of salvage logging on soil respiration of burnt forest have not been well studied and the combined effect of fire and logging were not fully explored. In turn, the difference of the effects caused by the fire and logging, and the effect difference of different logging methods (clear-cutting, selective-cutting, or no-cutting) have not been clarified.
In this study, a burnt forest dominated by Larix gmelinii (distributed in Daxing’anling and Xiaoxing 'anling mountains, also found in Russia and far east China) on permafrost were selected as the study area. The research aims to (1) compare the temporal changes in soil respiration and its components at different salvage logging intensities, (2) investigate the effects of fire disturbance and salvage logging interference on soil respiration over 13 years and (3) analyze the association of soil respiration between soil temperature and moisture, respectively.
2. Material and Methods
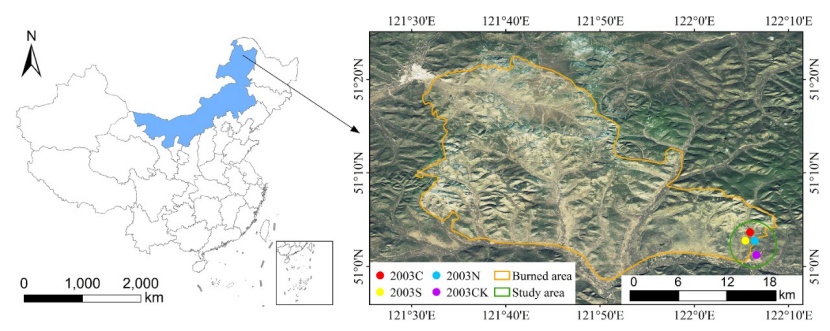
Figure 1: Location of study area und the burnt forest. The four experimental plots are located inside the green circle. / Abbildung 1: Lage des Untersuchungsgebiets und die Waldbrandfläche. Die vier Untersuchungsflächen liegen innerhalb des grünen Kreises.
2.1 Study Area
The study area located at the Kailaqi Forest Farm of the Genhe Forestry Bureau in Inner Mongolia, with the geographical position of east longitude:122°05΄21˝ ~ 122°04΄46˝, north latitude: 51°02΄46˝ ~ 51°01΄08˝ (Figure 1). The area is relatively flat, with elevations between 747 and 760 m. The area has a cold temperate and continental monsoon climate with an average annual temperature at -5.3 °C, the maximum daily temperature is 40 °C, and the minimum daily temperature is -58 °C. The average annual precipitation is 450 mm, which 12% as snowfall. The study area is located in a high latitude permafrost zone. Soil type is boralfs, which has a thin soil layer and a large number of gravel. Principal tree species are Larix gmelinii (Larix gmelinii (Rupr.) Kuzen.) and Betula platyphylla (Siberian silver birch) (Pan et al., 2013).

Figure 2: Field pictures of the four types of sample sites (photo taken in May 2018). 2003C is the clearcut, 2003S selective logged, 2003N unlogged and 2003CK the control site. At 2003C, small trees are not the original burnt wood, but newly re-generated trees. White rings show soil respiration measurement systems. / Abbildung 2: Photos der vier Probenstandorte (Foto vom Mai 2018). 2003C ist der Kahlschlag, 2003S wurde selektiv genutzt, 2003N nicht genutzt und 2003CK ist der Kontrollstandort. Bei2003C sind die kleine Bäume nach dem Brand gewachsen und nicht dieursprünglich verbrannten Bäume. Weiße Ringe zeigen Bodenatmungsmesssysteme an.
2.2 Sample plots
The chosen wildfire site was the Jinhe-Genhe forest fire, that occurred in 2003. The wildfire happened on May 5, 2003, burning for seven days and covering an area of 1330 km2 (east longitude: 121°29΄24˝ ~ 122°07΄52˝, north latitude: 51°22΄12˝ ~ 50°58΄26˝). Large-scale salvage logging management were conducted in the region in 2005, two years after the fire in 2003. There were three salvage logging methods implemented: clear-cutting, selective-cutting, and no-cutting. A control without burning and cutting was also established for comparison purpose. The three types of post-logging treatments were all light burned before cutting. Four sampling plots were set up in April 2016, ten years after the salvage logging occurred in 2005. The cutting intensity was 100% for the clear-cutting sample plot, 50% for the selective-cutting sample plot, 0% for the no-cutting sample plot and 0% for the control sample (unburnt-unlogged).
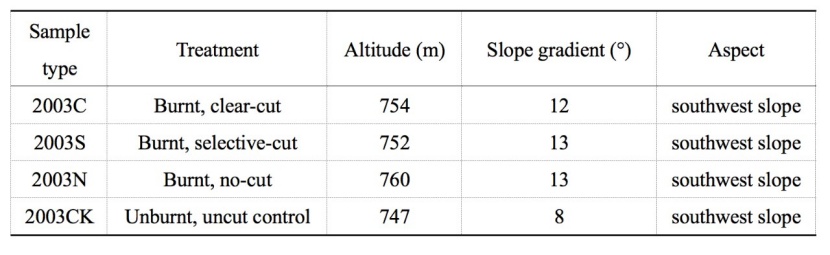
Table 1: Site characteristics of the four treatments (3 samplings sites per treatment). / Tabelle 1: Standorteigenschaften der vier Behandlungsvarianten (3 Probestellen pro Variante).
Three 30 m × 30 m observation sites were randomly distributed within each of the four treatments. The distance between the sample plots of the same treatment is about 10 m, the distance between the different treatments is about 500 m, giving a total of 12 observation sites (Figure 2, Table 1). An earlier field investigation in 2013 by Pan (2014) found that the 2003 forest fire affected soil respiration and that the soil respiration was still higher than the pre-fire level ten years after the fire. However, the effects of salvage logging (studied in the current study) were not measured in this earlier study.
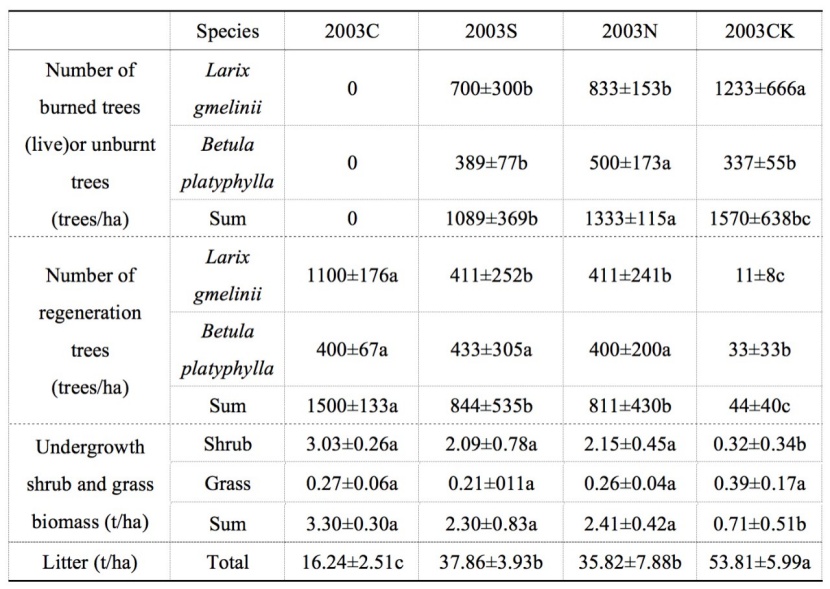
Table 2: The number of trees and undergrowth biomass of the 30 m×30 m sample plots. We show annual means with standard deviations. Letters (a, b, ab) show significant differences among salvage logging sample sites in the same line (Tukey’s HSD test). P-values were considered significant at α = 0.05. For the methods used for vegetation biomass and litter, we refer to the appendix. / Tabelle 2: Die Stammzahl und die Biomasse der Bodenvegetation und Streuschicht auf den 30 m × 30 m großen Plots. Die Werte sind jährliche Mittelwerte mit Standardabweichungen. Die Buchstaben (a, b, ab) zeigen signifikante Unterschiede zwischen den Probenahmestellen für die Bergungsprotokollierung in derselben Zeile (Tukey-HSD-Test). P-Werte wurden bei α = 0.05 als signifikant angesehen. Für die Methoden der Bodenvegetation und Streuschicth verweisen wir auf den Appendix.
An evaluation of species diversity found a total of 35 species of understory vegetation in the four experimental plots, belonging to 18 families and 32 genera. We measured factors (altitude, slope, slope direction) of each sample plot, and tree species and the number of regeneration trees (Table 2).
2.3 Method of soil respiration measurement
Soil respiration was measured by applying LI-8100 (LI-COR Inc.) soil respiration measurement system (Astiani et al., 2015). At the end of April 2016, three groups of soil respiration ring were installed in each sample plot (three rings used to measure total soil respiration, and three rings to measure heterotrophic soil respiration, each ring had a diameter of 20 cm and height of 13.5 cm). Each ring was inserted vertically into the soil, leaving 2 cm of the ring above the ground for soil respiration observation. Sets of soil respiration rings were randomly arranged in the sample plot in the form of an isosceles triangle, with a 10 m spacing between individual rings.
The measurement of the heterotrophic respiration used the trenching method (Figure 3). We selected a 1 m diameter circular area within 2 m of each soil respiration ring. Then in the circular area, a 30 cm wide trench was dug with a shovel until 0.4 m-0.6 m depth, based on earlier investigation, that roots in this area are mainly distributed in the soil layer of 0-10 cm, and at 40 cm the soil parent material is reached. Then the roots in the trench were cut off, but not removed, and then a solid plastic film was placed in the trench to prevent any roots from growing into the 1m-diameter circular soil column (Kuzyakov, 2006). Above ground vegetation on the undisturbed soil column was removed, and regularly cleaned up, in order to remove any autotrophic respiration from herbaceous vegetation. The measurement period spanned the growing season (May – October), for two years (2016, 2017). To observe the effect of freezing-thawing on soil respiration, the month of April was added to the soil respiration observation in 2017. Measurements were taken 2-3 times per month, from 8:00 a.m. - 12:00 a.m. on each sampling day.
Autotrophic respiration rate (RA) was calculated as:

Where RS represents total soil respiration rate (μmol m-2s-1), while RH is soil heterotrophic respiration rate. In equation (2) RA% represents the contribution ratio of soil autotrophic respiration relative to the total soil respiration (RS) (Han et al., 2018).

Figure 3: Pictures of soil rings for soil respiration measurements. A show the layout of heterotrophic soil respiration measurements using the trench method. B show a total soil respiration ring in the front. / Abbildung 3: Bilder der Bodenringe für Bodenatmungsmessungen. A zeigt einen Standort zur Messung der hetereotrophen Bodenatmung mit der Grabenmethode. B zeigt im Vordergrund einen Messring für die gesamte Bodenatmung.
2.4 Soil temperature and moisture observation
While observing soil respiration, we used a hand-held electron temperature probe (JM-222) to measure soil temperature at 10 cm depth (T10) at each experiment plot. A time-domain reflectometry sensor (TDR300, Spectrum, Aurora, USA) was used to measure soil moisture at a depth of 10 cm near each soil respiration ring (W10), and an averaged moisture value was obtained from all W10 measurements to represent each observation plot. The relationships between soil respiration rate (or its respiration components) and soil temperature or moisture were obtained based on the measured data; a regression model simulation of soil respiration was done as follows (Muñoz-Rojas et al., 2016).
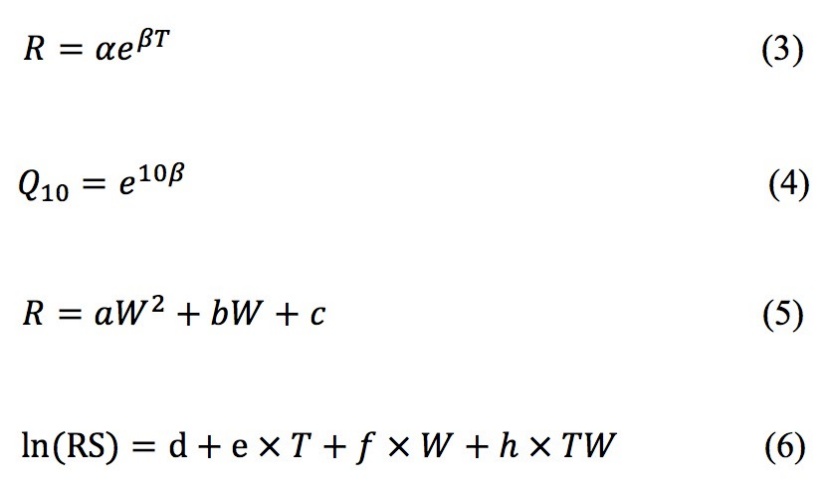
In equation (3) R is a random value of the RS, RA or RH rates, T represents the soil temperature (°C), α is the respiration rate at 0 °C, and β is a temperature response coefficient from which the temperature sensitivity coefficient Q10 can be obtained by equation (4), which indicates the sensitivity of soil respiration to temperature. The sensitivity index Q10 varies among different forest communities, different soil respiration components for the same community (Muñoz-Rojas et al., 2016). Equation (5) is a quadratic function of fitting R and soil moisture; a, b, and c are fitting constantly, and W is soil moisture (%). Equation (6) is a stepwise regression model, ln (RS) represents the natural logarithmic conversion of soil respiration rate, and d, e, f and h are fitting constantly (Wang et al., 2002).
3. Results
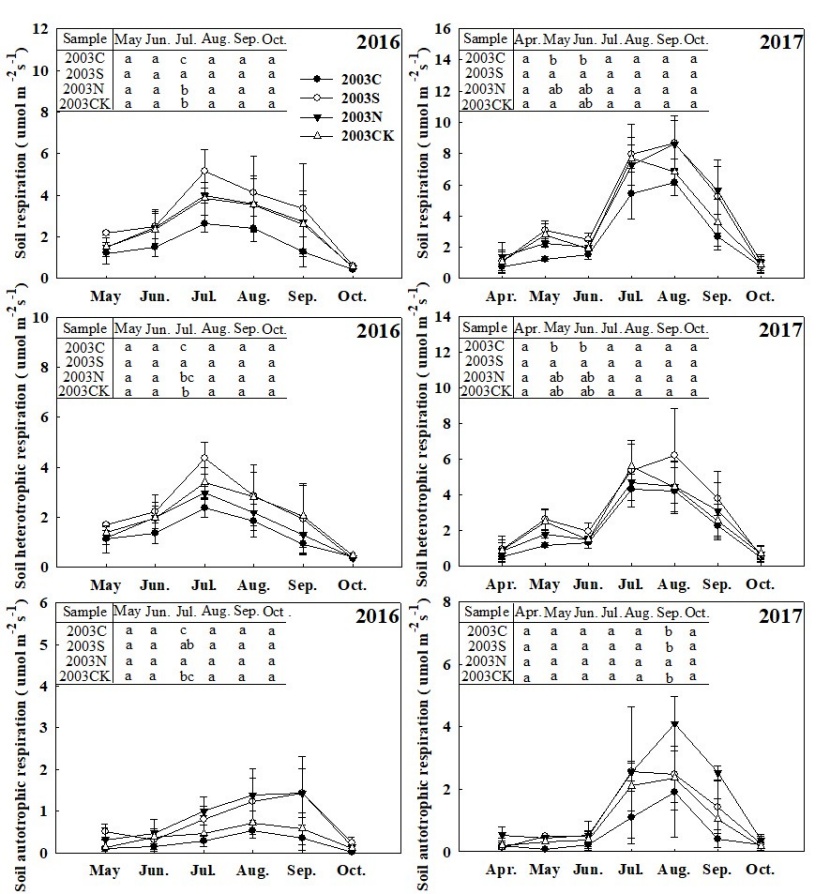
Figure 4: Monthly average soil respiration rate at four types of sample sites (see table 1). Letters (a, b, ab) show significant differences among sample sites in the same month and the same line (Tukey’s HSD test), p-values were considered significant at α = 0.05. / Abbildung 4: Monatliche durchschnittliche Bodenatmungsrate an vier Arten von Probenstandorten (siehe Tabelle 1). Die Buchstaben (a, b, ab) zeigen signifikante Unterschiede bei Stichprobenstellen im selben Monat und in derselben Zeile (Tukeys HSD-Test), P-Werte bei α = 0.05 wurden als signifikant angesehen.
3.1 Dynamics of soil respiration and its components in the growing season
Monthly mean RS, RH and RA at different salvage logging sample plots and control sample plot showed a single peak in the 2016 growing season. However, the peak months of each soil respiratory component were different (Figure 4), the first rising appeared in May – July (for RS and RH) and then decreased in August – October, but the peak of the RA component appeared in September. In July, the RS and RH of 2003S plot were significantly higher than that of 2003N, 2003C and 2003CK plot. The RA of 2003N plot was significantly higher than that of 2003C and 2003CK plot (P < 0.05). During the growing season of 2017, RS and RH in all plots showed a two-peak pattern, the first peak appearing in May and the second in August. RS and RH in 2003S plot were significantly higher than that in 2003C sample plot in May and June. The two-year data showed that RS and RH were higher in July and August (Figure 4).
The average annual-mean (or season-mean) RS appeared to be the largest at the selective-cutting plot 2003S, followed in turn by no-cutting 2003N, control 2003CK, and clear-cutting 2003C, the RS of 2003S plot is significantly higher than that of 2003C plot (Table 3). Partly similar to RS, the average annual-mean RH was the largest one at 2003S, followed by 2003CK, 2003N and 2003C. Moreover, RH in 2003S plot was significantly higher than that in 2003C plot, but the annual-mean RA showed a different pattern: largest at 2003N followed by 2003S, 2003CK and2003C, RA in 2003N plot was significantly higher than that in 2003C plot. For all respiration components, the burnt clear-cut produced the lowest rates compared to the other three plots, while the burnt selective-cut produced the most considerable rate (except for the RA). Based on two years of observation data, there are significant differences between 2003S plot and 2003C plot for both RS and RH, 2003N plot and 2003C plot for RA (Table 3). The differences between the two observation years may due to the inter-annual climate differences (Figure 5). Average air temperature in 2017 (9.5 °C) was higher than that in 2016 (6.04 °C). Air relative humidity is 66.08% and 65.07% in 2016 and 2017 respectively.
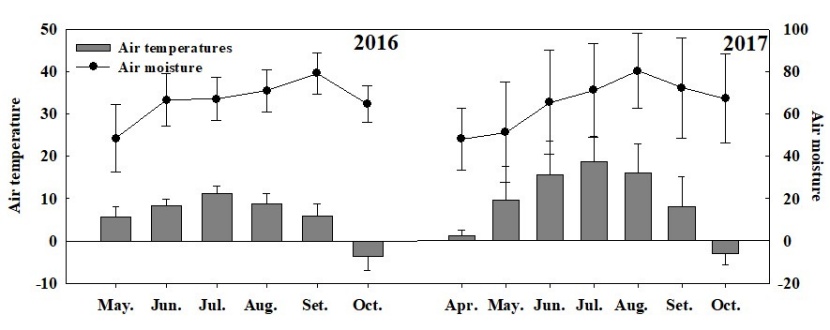
Figure 5: Monthly air temperature (°C) and air relative humidity (%) during the growing season in the research area. / Abbildung 5: Monatliche Lufttemperatur (°C) und relative Luftfeuchtigkeit (%) während der Vegetationsperiode im Forschungsgebiet.
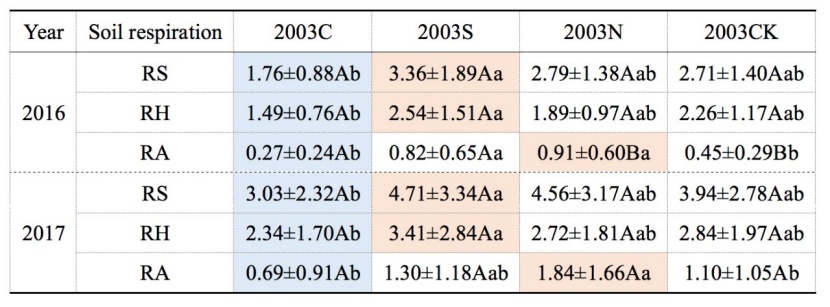
Table 3: Annual-mean and range of total soil respiration (RS), heterotrophic soil respiration (RH) and autotropic soil respiration (RA) (μmol m-2 s-1) in four types of sample sites. Values are annual means with standard deviations. Letters (a, b, ab) show significant differences among different sample sites in the same year and the same line. Letters (A, B) show significant differences in the same sample sites, and in different years (Tukey’s HSD test), P-values were considered significant at α = 0.05. The orange values are the highest rates, mostly in the 2003S and 2003N plots; the blue values are the lowest, usually appeared in the 2003C plot. / Tabelle 3: Jahresmittelwert und Bereich von gesamter Bodenatmung (RS), heterotropher Bodenatmung (RH) und autotropher Bodenatmung (RA) (μmol m-2 s-1) an vier Arten von Probenstellen. Die Werte sind Jahresmittelwerte mit Standardabweichungen. Die Buchstaben (a, b, ab) weisen erhebliche Unterschiede zwischen den verschiedenen Stichprobenstandorten im selben Jahr und in derselben Zeile auf. Die Buchstaben (A, B) zeigen signifikante Unterschiede zu denselben Stichprobenstandorten und in verschiedenen Jahren (Tukeys HSD-Test). Die P-Werte wurden bei α = 0.05 als signifikant angesehen. Die orange hervorgehobenen Werte zeigen die höchsten Raten hauptsächlich auf Plots mit selektiver bzw. ohne Nutzung. Die blau hervorgehobenen Werte sind die niedrigsten Raten auf Kahlschlagflächen.
3.2 Effects of different salvage logging on the contribution of soil respiration
Contribution of soil respiration varied from month to month during the growing season (Figure 6). Generally, the contribution ratio of RH% was higher (> 50% in most months) than the ratio RA% (< 50%), as observed at the four types of soil conditions. The contribution of RA at 2003N plot maintained a high level during the growing season (higher than other sites). In 2016 and 2017, the maximum RA% of 2003C plot appeared in August; while in 2003S plot, the maximum RA% appeared in July 2016 and September 2017; in 2003N and 2003CK plot, the maximum RA% appeared in September 2016 and August 2017 respectively. RH% had different trends in each site; in 2016, except the 2003C plot, the RH% maximum value of the other three sample sites appeared in May, and the minimum value appeared in September; in 2017, except the 2003S plot the RH% maximum value of the other three sample sites appeared in May, and the minimum value appeared in August. The average contribution of RA over the growing season was less than 35% for all four types of sample plots (Table 4). The averaged RA% at 2003N plot was greater than that of 2003CK control plot. The RA% after salvage logging curtailed (i.e. 2003C < 2003S < 2003N in Table 4), and the greater the cutting intensity, the smaller the contribution ratio of RA. By analyzing the observed data, there was a significant difference in RA% between 2003C and 2003S, 2003C and 2003N respectively.
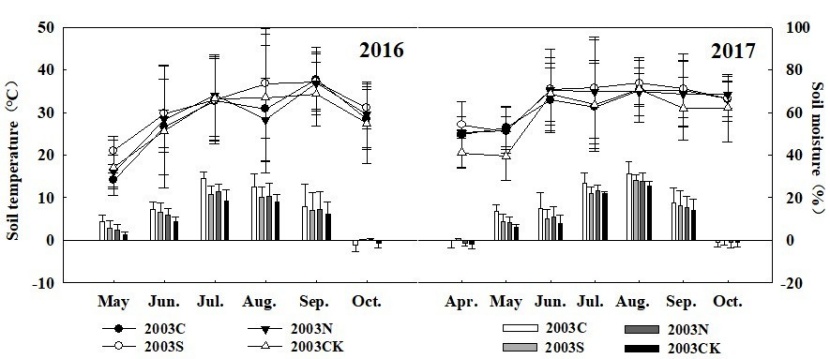
Figure 6: Monthly contribution ratio of soil respiration during growth season in the four types of sample sites. / Abbildung 6: Anteil der Bodenatmung pro Monat während der Vegetationsperiode in den vier Arten von Probenstandorten.
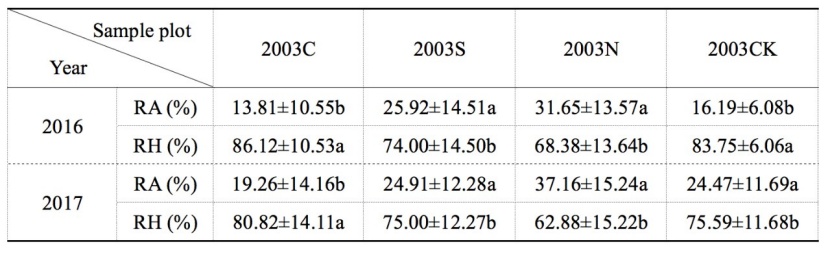
Table 4: Average contribution ratio of soil respiration RA and RH in four types of sample sites. Values are annual means with standard deviations. Letters (a, b, ab) show significant differences among salvage logging sample sites in the same line (Tukey’s HSD test). P-values were considered significant at α = 0.05. / Tabelle 4: Anteil der Bodenatmung RA und RH in vier Arten von Probenstandorten. Die Werte sind Jahresmittelwerte mit Standardabweichungen. Die Buchstaben (a, b, ab) zeigen signifikante Unterschiede zwischen den Probenahmestellen für die Bergungsprotokollierung in derselben Linie (Tukey-HSD-Test). Die P-Werte wurden bei α = 0.05 als signifikant angesehen.
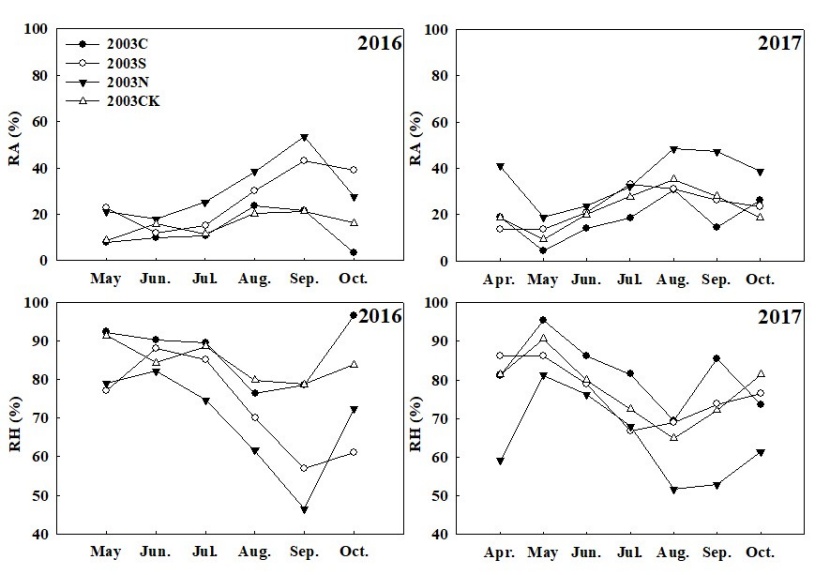
Figure 7: Monthly temperature and soil moisture at 10 cm depth, during the growing season, at four types of sample sites. / Abbildung 7: Monatliche Bodentemperatur und Bodenfeuchtigkeit in 10 cm Tiefe während der Vegetationsperiode an vier Arten von Probenstandorten.
3.3 Relationship between soil respiration and soil temperature-moisture
The monthly mean value of 10 cm soil temperature (T10) varied within the growing season (Figure 7). T10 reached its peak in July 2016 and August 2017 respectively. The growing-season averaged value of T10 in the salvage logging plots (2003C and 2003S) was larger than that of the unburnt and uncut control plot (2003CK). It can be seen that fire disturbance caused soil temperature to rise, and the logging disturbance caused the further soil temperature rise, and the greater the cutting intensity, the greater the soil temperature rose.
The monthly mean value of 10 cm soil moisture (W10) in the four types of sample plots had noticeable temporal variability within a season, and W10 in 2017 was higher than that in 2016 (Figure 7). The monthly mean value of W10 in 2016 peaked in September, with the minimum value in May. In 2017 it appeared to be the lowest and the highest in May and June respectively. During the two-year observation period, the minimum value of W10 appeared, probably due to the low precipitation in May, and because the vegetation in the region began to grow, the root system of vegetation absorbed a lot of soil water.
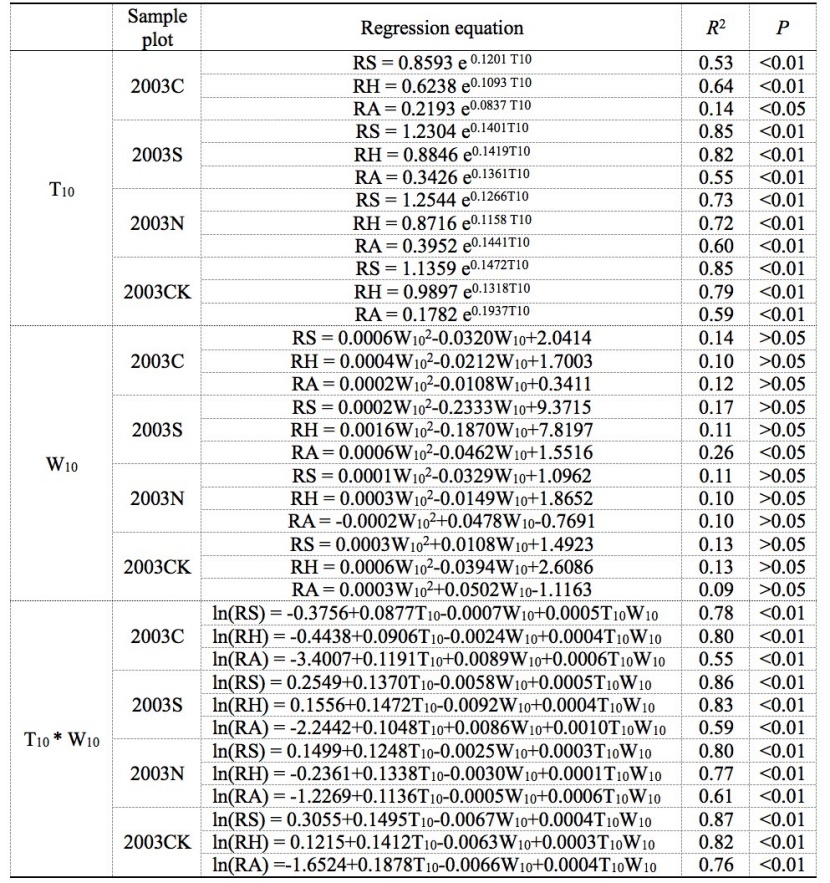
Table 5: Regression equations for soil respiration components with soil temperature (T10) and soil moisture (W10) for the four sample plots. R2 is coefficient of determination. / Tabelle 5: Regressionsgleichungen für Bodenatmungskomponenten mit Bodentemperatur (T10) und Bodenfeuchte (W10) auf den vier Probeparzellen. R2 ist das Bestimmtheitsmaß.
The exponential regression provided a good description of the correlation between soil respiration (or each soil respiration component) with soil temperature (Table 5), except for RA at the 2003C plot, the RS, RH and RA at three salvage-logging plots and the control plot enhanced with the increase of T10, and T10, which had a significant exponential relationship with soil respiration or its components (R2 > 0.50, P < 0.01 for two growing seasons). The RS and RH are well fitted with T10 at all four plots, but the RA was fitted relatively poor with T10 (R2 of RH is between 0.64 (P < 0.01) and 0.82 (P < 0.01); R2 of RA is between 0.13 (P > 0.05) and 0.59 (P < 0.01)). The correlation between RS and T10 in the burnt no-cut plot after 13 years of the fire disturbance was lower than that in the unburnt and uncut control plot (2003N R2 = 0.73 vs. 2003CK R2 = 0.85). Among the four types of sample plots, the 2003S plot had the highest fitting for RS or RH with T10 (R2 = 0.85; R2 = 0.83), the 2003CK plot had the maximum fitting for RA (R2 = 0.59), and the 2003C plots had the lowest fitting for RS, RH, or RA (R2 = 0.53; R2 = 0.64; R2 = 0.17). It can be seen that both the fire and harvest of fired trees affected the fitting of RS and soil temperature, making the correlation intensity varying.
The unitary quadratic regression model obtained to fit soil moisture W10, and soil respiration components (Table 5) gave low coefficients of determination. There were no significant correlations between soil respiration (or its components) and W10 at the four types of sample sites (R2 < 0.2 for most cases in Table 5, and P > 0.05), except for the 2003S sites RA and W10 (R2 = 0.26, P < 0.05).
The interaction between soil temperature and soil moisture (as treated by the term of W10×T10) and fitting degree with soil respiration rate and its components in the four sample sites were more suitable than single temperature factor or humidity factor; all regressions using equation (6) showed an extremely significant positive correlation (R2 < 0.55; P < 0.01). Except for the 2003S plot RH, the fitting degree of soil respiration or its components and (W10×T10) in the three types of burned wood management sample plot was lower than that in the 2003CK plot. The fitting degree was different in different types of burned wood management sample plot.
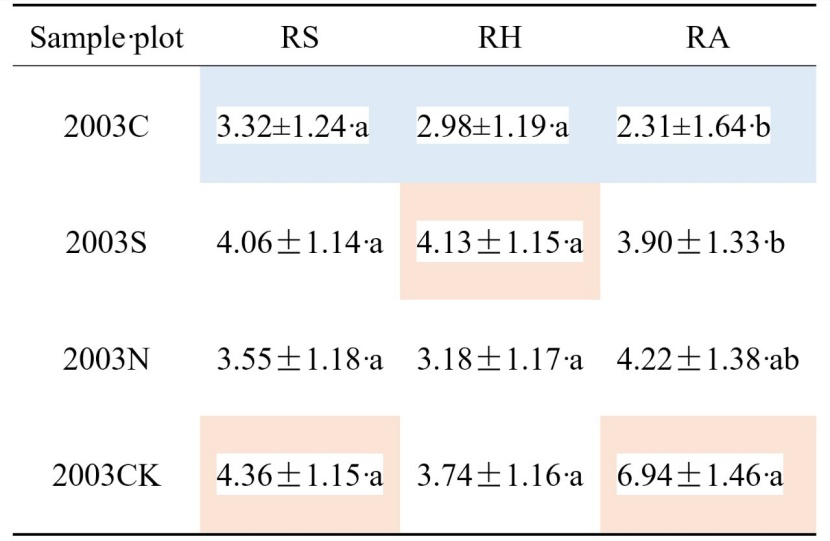
Table 6: Soil respiration component Q10 values in the growing season at four types of sample sites. Highlighted blue indicated the lowest and orange the highest values, respectively. Values are annual means with standard deviations. Letters (a, b, ab) show significant differences among salvage logging sample sites in the same line (Tukey’s HSD test). P-values were considered significant at α = 0.05. / Tabelle 6: Q10-Werte der Bodenatmungskomponenten in der Vegetationsperiode an vier Arten von Probenstandorten. Blau hervorgehoben zeigt die niedrigsten und orange die höchsten Werte an. Die Werte sind Jahresmittelwerte mit Standardabweichungen. Die Buchstaben (a, b, ab) zeigen signifikante Unterschiede zwischen den Probenahmestellen in derselben Linie (Tukey-HSD-Test). Die P-Werte wurden bei α = 0.05 als signifikant angesehen.
The three salvage-logging plots temperature sensitivity index Q10 for RS and RA were lower than that in 2003CK plot (Table 6). Among the four types of sample plots, the season-average value of Q10 for RS was the highest at 2003CK, showing a sensitivity trend as 2003CK > 2003S > 2003N > 2003C (Table 6); the average value of Q10 for RH was the largest at 2003S, showing a trend of 2003S > 2003CK > 2003N > 2003C; the Q10 for RA was the largest at 2003CK, showing a trend of 2003CK > 2003N > 2003S > 2003C. These trends indicated that soil respiration rates became less sensitive to soil temperature after the fire and logging interferences, which is consistent with what had happened in the respiration-temperature correlation described above. On the other hand, a comparison of Q10 across respiration components showed that the Q10 values for RA in 2003CK and 2003N plots were larger than for RS or RH (showing RA-Q10 > RS-Q10 > RH-Q10); the Q10 values for RH in 2003S were larger than for RS or RA (RH-Q10 > RS-Q10 > RA-Q10), the Q10 values for RS in 2003C were larger than for RH or RA (RS-Q10 > RH-Q10 > RA-Q10).
These patterns’ correlation and sensitivity showed that fire disturbance reduced the sensitivity of soil respiration and its components to soil temperature; and salvage logging also reduced the sensitivity, with the clear-cutting mostly reduced the sensitivity of soil autotrophic (P < 0.05). While Q10 in selective-cutting 2003S plot differed a little (only reduced from 4.36 to 4.05): less decreased in 2003N or 2003C for RS. However, Q10 in the selective-cutting 2003S plot became larger than in the 2003CK for RH (from 3.74 to 4.13).
4. Discussion
4.1 Fire disturbance on soil respiration and its components
The study indicated that the effects of fire disturbance on soil respiration varied with site location or condition for a given region. Hu et al. (2017) measured the effects of different fire intensity on soil respiration with the Larix gmelinii forest in another area of the same Daxing’anling mountains, where the fire broke out in 2006, with soil respiration monitored from 2010 to 2012. They found that both the soil respiration rate and soil autotrophic respiration in the burned sample plots were lower than those in the unburned plot. This finding is different from the results obtained in the study (the soil respiration and autotrophic respiration increased). Their research area located in the southeastern foothill of the Daxing’anling mountains, which was a temperate continental monsoon climate with no permafrost, and in which herbs and shrubs were abundant compared with the research area (northern Daxing’anling, permafrost), and the two study areas were 210 km apart. Fire changed soil structure and properties, which affected survival quantity and living environment of microorganisms (Holden et al., 2015), resulting in the RH of burnt sample plots, after 13 years since the fire event, being smaller than the unburnt control sample plot (i.e. smaller at 2003N than at 2003CK). At the same time, the fire changed the growth environment and mechanism of trees, which affected the root respiration of fired trees and understory vegetation (Bryanin and Makoto 2017). After 13 years from fire disturbance, the vegetation restoration was good, the number of tree regeneration increased, and the biomass of understory vegetation was higher, which promoted the soil autotrophic respiration (Table 2). RA at the burned plots (2003N) became higher than that of the unburnt control plot (2003CK).
The contribution ratio of heterotrophic respiration was higher than the ratio of autotrophic respiration (RH% > RA%), which is similar to the previous studies (e.g. Heim et al., 2015; Lin et al., 1999). The RA% in the burnt plots was more significant than that in the no-fire plot. However, the salvage logging intensity decreased it: the higher the cutting intensity, the smaller the ratio. The reason may because that fire reduced the amount and activity of soil microorganisms (Wang et al., 2012), and changed the growth environment and growth mechanism at the same time, thus caused the RH to weaken and RA to increase.
Potential response of soil respiration to changes in environmental factors can be separated into the response of autotrophic and heterotrophic respiration, as their responses to soil temperature may vary (Archmiller and Samuelson, 2016). At research sites, the regression fitting strength of soil respiration (or its components) with soil temperature is high, similar to the results of Jian et al. (2019). The study provided a useful addition of the respiration-temperature relation for cold, permafrost regions, and confirmed the sensitive response of soil autotrophic respiration to temperature change. As found by others (Lu et al., 2010, Epron et al., 2001), the autotrophic sensitivity Q10 value (6.94) of the unburnt and uncut sample plot was higher than the RH’s Q10 value (3.74), supporting that autotrophic respiration is more sensitive than heterotrophic respiration in the Larix gmelinii forest in the cold temperate zone of a permafrost region. The Q10 values for all RS, RA and RH at the burnt without cut plot were lower than that of the unburnt and uncut control plot, which suggested that the fire disturbance reduced the sensitivity of soil respiration to temperature, probably because burnt plots had thinner litter layer and weaker decomposition ability (Davidson et al., 2006).
4.2 Difference of soil respiration and its components among different salvage-logging management method
The results indicated a contrasting effect between the selective-cutting (promoting soil respiration) and the clear-cutting (lowering soil respiration). The findings of positive effect in selective-cutting (increasing soil respiration rates) observation here is similar to Concilio et al. (2005) for burnt and thinned mixed-conifer and hardwood forests. Some other studies could support the negative or lowering effect of clear-cutting at the site, e.g., López-Serrano et al. (2016) for a Mediterranean mixed forest. However, some other studies have shown contrarily (e.g. Parro et al., 2019) on burned and clear-cutting plots compared to burned and uncleared plots, showing no differences in respiration rates. Ma et al. (2013) suggested that felling had no significant effects on cumulative annual soil respiration releases, mainly because root respiration usually stopped shortly after tree harvesting, resulting in a sudden decrease in soil autotrophic respiration. However, the decomposed dead roots increased the number and activity of microorganisms, which could counteract the decline in root respiration. At the same time, the newly increased growth of herbaceous and other plant roots within the sample plots may offset the soil autotrophic respiration reduction.
Salvage logging affected the further soil respiration and changed the effects or patterns which were initially formulated by the fire event. Moderate logging (selective-cutting 2003S) was benefited to the vegetation renewal and soil microbial activities in the fired area, increasing the RH against other two logging practices (i.e. giving an RH rate pattern of 2003S > 2003N > 2003C). It is observed that the minimums of RS, RH and RA were all found in the clear-cutting (2003C) plot investigated as the high-intensity cutting has a lower effect on soil respiration and its components, because the high-intensity cutting of burned wood changed the aboveground vegetation composition and biomass (Takada et al., 2015). Most aboveground vegetation was new after the burned wood cleared, the RA rate became weak, and the underground microbial amount was reduced, which led to the soil respiration components at the 2003C plot to became lower compared to the other plots.
The changes in contribution ratios could also be explained by the changes in soil temperature and vegetation amount. Nakane et al. (1996) suggested that the root respiration was strictly related to the amount of vegetation on the ground, accounting for about 30% of total respiration (which is comparable to the ratios of the sites). Lin et al. (1999) found that the increase in temperature would increase the contribution rate of soil autotrophic respiration. The soil temperature at depth 10 cm at salvage-logging plots was higher than at the no-fire no-cut control plot, which was caused by the changed tree growth environment due to fire. As a result, the vegetation root respiration rate (i.e. soil autotrophic respiration contribution ratio) increased after the fire. However, the salvage-logging management reduced the biomass of trees and changed the forest canopy density. The higher the cutting intensity, the smaller the proportion of soil autotrophic respiration would be at the fired plots (as shown by 2003N > 2003S > 2003C).
Both fire and logging increase soil temperature, and the incremental temperature at the cut plot after a fire is higher than that at the burnt no-cut plot (Concilio et al., 2005). The study provided another confirmation. The sensitivity also differed from salvage logging methods, as shown a greater Q10 at the 2003S plot than at 2003N or 2003C plot. Cutting has a substantial impact on the biomass of soil roots. After ten years from the salvage-logging management, the clear cutting (2003C) reduced the number of mature trees. Therefore, the sensitivity of autotrophic respiration to temperature may be related to the number of roots biomass of mature trees. Soil moisture may not be a key factor affecting the soil respiration of the Larix gmelinii forest in the permafrost region, In terms of the experiment, there were lack of no noticeable changes in soil moisture among the three salvage-logging management plots and the unburnt and uncut control plot. Holden et al. (2015) found no significant differences in soil moisture between fired and unfired plots, which was consistent with the research findings.
5. Conclusions
After thirteen years fire disturbance and ten years salvage-logging management at a cold and permafrost area in the Daxing'anling mountains, the impact of fire and logging on soil respiration of the studied area still existed. Forest fire increased soil autotrophic respiration but decreasing soil heterotrophic respiration, resulting in a slightly increased total soil respiration compared to the unburnt and uncut control plot. This research will report in the first time that: the selective-cutting of burnt tree enhanced the soil respiration and its components, producing most considerable respiration rates at the selective-cutting plot among different cutting intensities. In contrast, clear-cutting weakened soil respiration and its components produced the lowest respiration rates. The proportion of heterotrophic soil respiration to soil respiration was much more significant than the ratio of autotrophic respiration. The fire disturbance increased the proportion of soil autotrophic respiration, but the salvage logging reduced it, and the higher the cutting intensity, the lower the contribution ratio. The logging of burnt trees increased soil temperature, and the greater the cutting intensity, the higher the temperature would be. Although soil respiration and its components were positively correlated to soil temperature in all plots, both the fire and logging interferences had reduced the sensitivity of soil respiration to soil temperature. There were no significant correlations between soil respiration and soil moisture in the studied forest. The results suggested clearly that the direction (positive or negative) and extent of the effects on soil respiration could be quite different between fire disturbance and post-fire logging, and the effects can differ significantly from logging methods. Based on current analysis of soil respiration data, we prefer not to conduct burnt wood logging. However, soil respiration is the only index of soil and forest ecosystem, scientifically sound management policy for fired forest may not be recommended without considering all ecological and forestry factors. More studies and experiments are required.
Author’s contributions
M. Zhou and Z. Wang initiated the research plan; Z. Wang synthesized the field work and results; Z. Wang and H. Yao wrote the manuscript; D. Wang conducted some statistical work and helped writing the manuscript. P. Zhao helped field work; A. James helped writing the manuscript.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (41563006), Inner Mongolia autonomous region doctoral research and innovation project in 2018, Inner Mongolia Agricultural University (B2018111945). Comments from David Aldred improved the paper writing. Support was provided by the CFERN&TECHNO Award Fund for Outstanding Academic Achievements. Comments and suggestions from peer reviewers are appreciated.
Conflict of interest
None declared.
Appendix
Vegetation investigation
Identifying regenerated trees was based on the tree height. The forest fire broke out in 2003 and a vegetation survey was conducted in 2016. The regenerated trees were maximum 13 years old and the height of regeneration of Larix gmelinii and Betula platyphylla was no more than 2.5 m, which made it easy to identify them. The post-fire logging did not prefer particular tree species, but removed trees according to the degree of burning.
Five 5 m× 5 m quadrats were set at the four corners and the center of each sample plot respectively to investigate shrub indicators. Five 1 m×1 m quadrats were randomly set in each shrub quadrat to investigate herb indicators. Species, height, number of plant clusters and coverage were investigated in the shrub and herb quadrats. In order to preserve the integrity of the plot, the shrub was sampled by the standard branch method. According to the survey data of each shrub in the sample plot, the plants in whole sample plot were divided into three grades respectively according to the arithmetic mean of three indexes - the plant height, ground diameter and the sum of height and ground diameter, then three shrubs were selected for each grade (Liu, 1994). The biomass of aboveground shrubs was determined by Monsi layer cutting method (Monsi, 2005). The whole harvest method will be applied, if a shrub were not meet the selection criteria for the standard wood. Herbs were collected by total harvest method. Taking three litter quadrats of 0.2 m × 0.2 m randomly and setting in each plot, including deciduous grass and branches with diameters less than 2 cm, excepting for fallen wood and branches with diameters greater than 2 cm. We measured the fresh mass after sampling shrubs, herbs and litter, then brought the material back to the laboratory and dried at 80 °C until the mass was constant and weighed again. The biomass per unit area and litter amount of each shrub and herb were estimated according to the quadrat mean value.
References
Aaltonen, H., Palviainen, M., Zhou, X., Köster, Egle, Berninger, F., Pumpanen, J. 2019. Temperature sensitivity of soil organic matter decomposition after forest fire in Canadian permafrost region. Journal of Environmental Management, 241(7-1), 637-644. doi: 10.1016/j.jenvman.2019.02.130.
Archmiller, A., Samuelson, L. 2016. Partitioning longleaf pine soil respiration into its heterotrophic and autotrophic components through root exclusion. Forests, 7(2), 39. doi: 10.3390/f7020039.
Astiani, D., Hatta, M., Fifian, F. 2015. Soil CO2 respiration along annual crops or land-cover type gradients on West Kalimantan degraded peatland forest. Procedia Environmental Sciences, 28, 132-141. doi: 10.1016/j.proenv.2015.07.019.
Bryanin, S. V., Makoto, K. 2017. Fire-derived charcoal affects fine root vitality in a post-fire Gmelin larch forest: field evidence. Plant and soil, 416(1-2), 409-418. doi: 10.1007/s11104-017-3217-x.
Concilio, A., Ma, S., Li, Q., Lemoine, J., Chen, J., North, M. Jensen, R. 2005. Soil respiration response to prescribed burning and thinning in mixed-conifer and hardwood forests. Canadian journal of forest research, 35(7), 1581-1591. doi: 10.1139/x05-091.
Davidson E A, Richardson A D, Savage K E, Hollinger D Y . A distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce-dominated forest. Global Change Biology, 2006, 12(2):230-239. doi: 10.1111/j.1365-2486.2005.01062.x.
Eom, J. Y., Jeong, S. H., Chun, J. H., Lee, J. H., Lee, J. S. 2018. Long-term characteristics of soil respiration in a Korean cool-temperate deciduous forest in a monsoon climate. Animal cells and systems the official publication of the Zoological Society of Korea, 22(2), 100-108. doi: 10.1080/19768354.2018.1433234.
Epron, D., Le Dantec, V., Dufrene, E., Granier, A. 2001. Seasonal dynamics of soil carbon dioxide efflux and simulated rhizosphere respiration in a beech forest. Tree Physiology, 21(2-3), 145-152. doi: 10.1093/treephys/21.2-3.145.
Francos, M., Pereira, P., Mataix-Solera, J., Arcenegui, V., Alcañiz, M., Úbeda, X. 2018. How clear-cutting affects fire severity and soil properties in a Mediterranean ecosystem. Journal of environmental management, 206, 625-632. doi: 10.1016/j.jenvman.2017.11. 011.
Garcia-Carmona, M., Arcenegui, V., Garcia-Orenes, F., Mataix-Solera, J. 2020. The role of mosses in soil stability, fertility and microbiology six years after a post-fire salvage logging management. Journal of Environmental Management, 262(5.15), 110287.1-110287.8.doi: 10.1016/j.jenvman. 2020.110287.
Haeussler, S., Bergeron, Y. 2004. Range of variability in boreal aspen plant communities after wildfire and clear-cutting. Canadian Journal of Forest Research, 34(2), 274-288. doi: 10.1139/x03-274.
Han, M., Jin, G. 2018. Seasonal variations of Q10 soil respiration and its components in the temperate forest ecosystems, northeastern china. European Journal of Soil Biology, 85, 36-42. doi: 10.1016/j.ejsobi.2018.01.001.
Heim, B. C., Seiler, J. R., Strahm, B. D. 2015. Root nonstructural carbohydrates and their relationship with autotrophic respiration of loblolly pine (Pinus taeda L.). Communications in soil science and plant analysis, 46(7), 888-896. doi: 10.1080/00103624.2015.1011752.
Holden, S. R., Berhe, A. A., Treseder, K. K. 2015. Decreases in soil moisture and organic matter quality suppress microbial decomposition following a boreal forest fire. Soil Biology and Biochemistry, 87, 1-9. doi: 10.1016/j.soilbio.2015.04.005.
Hu, T., Sun, L., Hu, H., Weise, D. R., Guo, F. 2017. Soil respiration of the dahurian larch (larix gmelinii) forest and the response to fire disturbance in da xing'an mountains, china. entific Reports, 7(1), 2967. dio: 10.1038/s41598-017-03325-4.
Hubbard, R. M., Ryan, M. G., Elder, K., Rhoades, C. C. 2005. Seasonal patterns in soil surface CO2 flux under snow cover in 50 and 300 year old subalpine forests. Biogeochemistry, 73(1), 93-107. doi: 10.1007/s10533-004-1990-0.
Jian, S., Zhen, L., Yuan, Z., Tao, Y., Zehao, S., Shilong, P. 2019. Effects of wildfire on soil respiration and its heterotrophic and autotrophic components in a montane coniferous forest. Journal of Plant Ecology, 12(2):336-345. doi:10.1093/jpe/rty031.
Kelly, R., Genet, H., McGuire, A. Hu, F. S. 2016. Palaeodata-informed modelling of large carbon losses from recent burning of boreal forests. Nature Climate Change, 6(1): 79-82. doi: /10.1038/nclimate2832.
Kim, J. S., Lim, S. H., Joo, S. J., Shim, J. K., Yang, K. C. 2014. Characteristics of soil respiration in Pinus densiflora stand undergoing secondary succession by fire-induced forest disturbance. Journal of Ecology and Environment, 37(3), 113-122. doi: 10.5141/ecoenv.2014.014.
Köster E, Köster K, Berninger F, et al. 2018. Changes in fluxes of carbon dioxide and methane caused by fire in Siberian boreal forest with continuous permafrost. Journal of environmental management, 228: 405-415. doi: 10.1016/j.jenvman.2018.09.051.
Kuzyakov, Y. 2006. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biology and Biochemistry, 38(3), 425-448. doi: 10.1016/j.soilbio.2005.08.020.
Lafleur, B., Zouaoui, S., Fenton, N. J., Drapeau, P., Bergeron, Y. 2016. Short-term response of Cladonia lichen communities to logging and fire in boreal forests. Forest Ecology and Management, 372, 44-52. doi: 10.1016/j.foreco.2016.04.007.
Lin, G., Ehleringer, J. R., Rygiewicz, P. T., Johnson, M. G., Tingey, D. T. 1999. Elevated CO2 and temperature impacts on different components of soil CO2 efflux in Douglas‐fir terracosms. Global Change Biology, 5(2), 157-168. doi: 10.1046/j.1365-2486.1999.00211.x.
Liu C Q.1994. Study on the measurement technology of shrub plants. Acta Prataculturae Sinica, 003(004):61-65. doi: cnki: sun: cyxb.0.1994-04-009. (in Chinese)
López-Serrano, F. R., Rubio, E., Dadi, T., Moya, D., Andrés-Abellán, M., García-Morote, F. A., Martínez-García, E. 2016. Influences of recovery from wildfire and thinning on soil respiration of a Mediterranean mixed forest. Science of The Total Environment, 573, 1217-1231. doi: 10.1016/j.scitotenv.2016.03.242.
Lu, B., Wang, S. H., Mao, Z. J., Sun, T., Jia, G. M., Jin, S. B., Cheng, C. X. 2010. Soil respiration characteristics of four primary Korean pine communities in growing season at Xiaoxing’an Mountain, China. Acta Ecologica Sinica, 30, 4065-4074. doi: cnki:sun:stxb.0.2010-15-013. (in Chinese)
Ma, Y., Geng, Y., Huang, Y., Shi, Y., Niklaus, P. A., Schmid, B., He, J. S. 2013. Effect of clear-cutting silviculture on soil respiration in a subtropical forest of China. Journal of Plant Ecology,6(5), 335-348. doi: 10.1093/jpe/rtt038.
Martínez-García, E., López-Serrano, F.R., Dadi, T., García-Morote, F.A., Andrés-Abellán, M., Pumpanen, J. 2017. Medium-term dynamics of soil respiration in a mediterranean mountain ecosystem: the effects of burn severity, post-fire burnt-wood management, and slope-aspect. Agricultural & Forest Meteorology, 233(Complete), 195-208.doi: 10.1016/j.agrformet.2016.11.192.
Molchanov, A. G., Kurbatova, Y. A., Olchev, A. V. 2017. Effect of clear-cutting on soil CO2 emission. Biology Bulletin, 44(2), 218-223. doi: 10.1134/S1062359016060121.
Monsi, M., Saeki, T. 2005. On the factor light in plant communities and its importance for matter production (an English translation of Monsi and Saeki (1953)). Annals of Botany, (95):549-567. doi: 10.1093/aob/mci052.
Muñoz-Rojas, M., Lewandrowski, W., Erickson, T. E., Dixon, K. W., Merritt, D. J. 2016. Soil respiration dynamics in fire affected semi-arid ecosystems: Effects of vegetation type and environmental factors. Science of the Total Environment, 572, 1385-1394. doi: 10.1016/j.scitotenv. 2016.02.086.
Nakane, K., Kohno, T., Horikoshi, T. 1996. Root respiration rate before and just after clear‐felling in a mature, deciduous, broad‐leaved forest. Ecological Research,11(2), 111-119. doi: 10.1007/ BF02347678.
Pan, L. 2014. Study on soil greenhouse gases effluxes in the burned areas of coniferous forest in cold temperate zone. Doctoral dissertation, PhD Dissertation. Inner Mongolia Agricultural University. Hohhot, China. (in Chinese)
Pan, L., Mei, Z., PengWu, Z., JiangSheng, W., QingHai, W., Xiang, C., Kezhen, Q. 2013. Soil respiration and its relationships with hydrothermic factors in the burned areas of daxingan mountain. Chinese Journal of Ecology,32(12),3305-3311. doi: 10.13292/j.1000-4890.2013.0505. (in Chinese)
Parro, K., Köster, K., Jõgiste, K., Seglinš, K., Sims, A., Stanturf, J. A., Metslaid, M. 2019. Impact of post-fire management on soil respiration, carbon and nitrogen content in a managed hemiboreal forest. Journal of environmental management, 233, 371-377. dio: 10.1016/j.jenvman.2018.12.050.
Serrano-Ortiz, P., S.Marañón-Jiménez, Reverter, B. R., E.P. Sánchez-Cañete, Castro, J. Zamora, R. 2011. Post-fire salvage logging reduces carbon sequestration in mediterranean coniferous forest. Forest Ecology and Management,262(12), 2287-2296. doi:10.1016/j.foreco.2011.08.023.
Song X, Wang G, Hu Z, et al.2018. Boreal forest soil CO2 and CH4 fluxes following fire and their responses to experimental warming and drying. Science of the total environment, 644: 862-872.dio: 10.1016/j.scitotenv.2018.07.014.
Stevens, A., Van wesemael, B. 2008. Soil organic carbon stock in the Belgian Ardennes as affected by afforestation and deforestation from 1868 to 2005. Forest Ecology and Management, 256(8), 1527-1539. doi: 10.1016/j.foreco.2008.06.041.
Takada, M., Yamada, T., Shamsudin, I., Okuda, T. 2015. Spatial variation in soil respiration in relation to a logging road in an upper tropical hill forest in Peninsular Malaysia. Tropics, 24(1), 1-9. doi: 10.3759/tropics.24.1.
Tang, X., Pei, X., Lei, N., Luo, X., Liang, J. 2020. Global patterns of soil autotrophic respiration and its relation to climate, soil and vegetation characteristics. Geoderma, 369, 114339. doi: 10.1016/j.geoderma.2020.114339.
Thiffault, E., Bélanger, Nicolas, Paré, David, Munson, A. D. 2007. How do forest harvesting methods compare with wildfire? a case study of soil chemistry and tree nutrition in the boreal forest. Canadian Journal of Forest Research, 37(9), 1658-1668. doi: 10.1139/X07-031.
Wang C, Ben Bond‐Lamberty, Gower S T. 2002. Soil surface CO2 flux in a boreal black spruce fire chronosequence. Journal of Geophysical Research: Atmospheres, 107(D3). doi: 10.1029/2001 JD000861.
Wang, Q., Zhong, M., Wang, S. 2012. A meta-analysis on the response of microbial biomass, dissolved organic matter, respiration, and N mineralization in mineral soil to fire in forest ecosystems. Forest Ecology and Management, 271, 91-97. doi: 10.1016/j.foreco.2012.02.006.
Wittenberg, L., Wal, H. V. D., Keesstra, S., Tessler, N. 2020. Post-fire management treatment effects on soil properties and burned area restoration in a wildland-urban interface, Haifa Fire case study. The Science of the Total Environment, 716, 135190.1-135190.10. doi: 10.1016/j.scitotenv.2019.135190.



