Schlüsselbegriffe: natürliche Astreinigung, Allometrie, Allokationsmuster, Eigenschaften von Zweigen, Verteilung der Zweig, astfreies Holz
Abstract
Branch development is related to the formation of wood knots and affects timber quality and future market value of plantations and native forests alike. This study provides a theoretical basis and reference for knot-free timber production in a Mytilaria laosensis plantation. The branch diameter, length, angle, status (alive or dead), azimuth, and branch height under 12 m were measured for 60 dominant or co-dominant trees in response to five planting density treatments (625, 833, 1111, 1667, and 2500 stems ha-1). With increasing planting density from 625 to 2500 stems ha-1, the mean branch diameter and branch length decreased by 7.09% and 8.70% respectively. Dead and live branch angle decreased by 10.54% and 9.66% respectively, while number of living branches increased by 31.67% from 625 to 2500 stems ha-1. Analyzing the spatial distribution using branch azimuth, we discovered that branches pointing north were more abundant and had larger diameter, larger branch length, and higher branch angle than branches pointing south. Branch diameter, length, and number were positively related to the relative branch height. Our results underline that increasing the planting density from 625 to 2500 stems ha-1 considerably affect branch growth. Thus, optimizing planting density when establishing Mytilaria laosensis plantations according to the forest management targets and future pruning can improve the timber quality and future value.
Zusammenfassung
Die Zweigentwicklung hängt mit der Bildung von Astlöchern und Knoten zusammen, die sich wiederum auf die Holzqualität und den zukünftigen Marktwert eines Bestandes auswirken. Ziel dieser Studie war es, eine theoretische Grundlage und Referenz für astfreie Holzproduktion einer Mytilaria laosensis Plantage zu erarbeiten. Wir untersuchten den Astdurchmesser, die Astlänge, der Astwinkel, den Status (lebendig oder tot) der Äste, die Orientierung der Äste sowie die Asthöhe unter 12 m für 60 dominante oder kodominante Bäume in fünf Pflanzdichtevariante (625, 833, 1111, 1667 und 2500 Stämme pro Hektar). Mit zunehmender Pflanzdichte von 625 auf 2500 Stämme ha-1 nahm der mittlere Astdurchmesser und die Astlänge ab (-7.09 % bzw. -8.70 %). Der Winkel wurde zunehmend spitzer sowohl für lebende als auch tote Äste (-10.54 % bzw. -9.66 %) und die Anzahl der lebenden Äste wurde um 31.67 % größer mit zunehmender Pflanzdichte. Hinsichtlich der räumlichen Verteilung der Äste zeigt sich, dass Äste in nördlicher Richtung höhere Astdurchmesser und Astlänge und geringere Astwinkel aufwiesen als Äste in südlicher Richrung. Durchmesser, Länge und Anzahl der Äste standen in positivem Zusammenhang mit der relativen Asthöhe. Diese Ergebnisse legen nahe, dass eine Erhöhung der Pflanzdichte von 625 auf 2500 Stämme ha-1 das Zweigwachstum deutlich beeinflusst. Die Wahl einer geeigneten Pflanzdichte beim Anlegen von Mytilaria laosensis Plantagen entsprechend den waldbaulichen Zielen und der Möglichkeit eines Astschnitt kann daher die Holzqualität und den Marktwert erhöhen.
Introduction
Mytilaria laosensis Lecomte is a fast-growing, broad-leaved tree species that has straight stem form and growing in China’s South Subtropical Region (Guo et al. 2006). This species has been widely used for forestry transformation and timber production (Lin et al. 2000; Chen et al. 2012). High-quality and large-diameter M. laosensis timber is becoming increasingly popular on the markets. However, the low natural pruning capacity of M. laosensis results in the formation of many knots negatively affecting wood development and utilization.
Branches are a vital part of the canopy and support the leaves and reproductive structures. Tree vitality, growth, and stem form are determined by the distribution and size of the branches (Biging and Dobbertin 1992), which in turn affects tree growth and the forest stand volume (Wang et al. 2007). The quality and value of timber depend largely on the branches, as the diameter and number of branches along the stem are closely related to knot development (Hein 2008; Wang et al. 2016b). Dead branches, that are not shed, are gradually wrapped by the trunk cambium. Artificial pruning is a traditional forest management method critical for cultivating timber without knots, as it can reduce not only water and nutrient consumption (O’Hara 1991), but also the dead-knot defects of wood (Hein and Spiecker 2007). However, this method is time-consuming and costly. Therefore, studies of branch characteristics and distributions are critical to understand the growth characteristics and natural pruning. Stand density can control branch development and reduce costs for pruning and improve the quality of timber (Kellomäki et al. 1989).
Numerous factors influence branch development, including stand density (Mäkinen 1999a; Neilsen and Gerrand 1999; Alcorn et al. 2007), water and nutrient allocation to branches (Mäkinen et al. 2001), genotype (Alcorn et al. 2007; Gort et al. 2010), and other factors. For timber species, the lower part of stem below 6-8 m accounts for 90% of the future tree value (Kint et al. 2010). Thus, branch development in this region is essential for the early growth of trees and later utilization of the wood (Wang et al. 2018). Density control is an integral part of plantation management. Branch death and the speed of wound healing can be accelerated by high-density planting (Johansson 1992; Mäkinen 1999a). Although wide spacing is beneficial for the development of large-diameter wood, it can prolong branch death (Mäkinen 2002).
We previously investigated the effect of different planting densities on branch development in ‘standard trees’ of a six-year-old M. laosensis plantation (Wang et al. 2019). The definition of standard trees are trees with average diameter and tree height within the forest. Standard trees are representative for an unthinned forest. Selective target tree management and thinning has gained more attention from national and international forest researchers (e.g. De Freitas, 2004; David C, 2007; Liao SX, 2009). Dominant or co-dominant trees are the main crop trees for future harvesting, and suppressed trees may be removed by thinning. This approach can result in high-quality, large-diameter wood for the market, support sustainable forest management, and maximize the stability of forest ecosystems. We measured branch number, status (alive-dead), branch diameter, branch length, branch angle, azimuth, and height of first-order branches under 12 m of 60 dominant or co-dominant trees M. laosensis trees at different planting densities. We then tested the effect of five planting densities on branch development in this eight-year-old M. laosensis plantation. The results of the present study can provide an appropriate planting density for producing high-quality M. laosensis timber and reduce costs for artificial pruning.
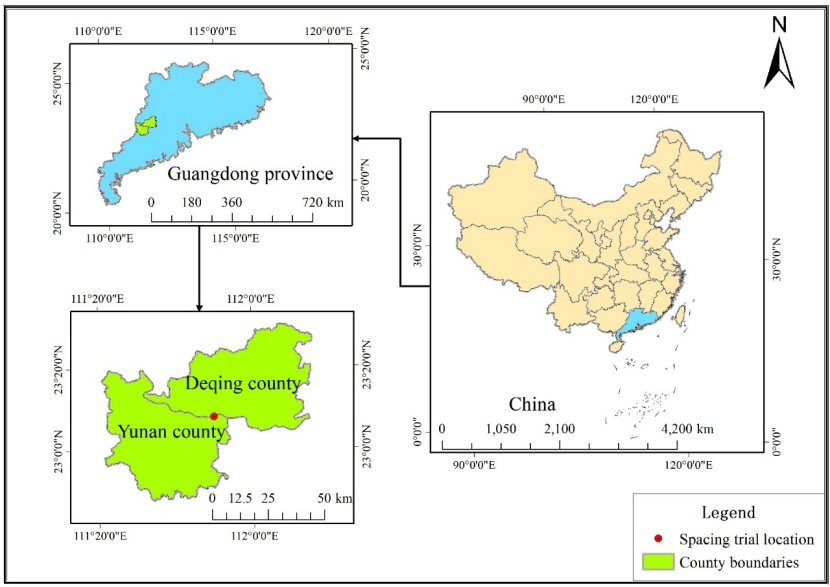
Figure 1: Location of M. laosensis spacing trial in Yunan County, Guangdong Province. / Abbildung 1: Die Lage des M. laosensis Pflanzdichteversuchs im Bezirk Yunan der Provinz Guangdong
Experimental Site
The spacing trial was located at the SanKeng experimental base, XiJiang Forest Farm, Yunfu City, Guangdong Province, China (23°07΄N, 111°51΄E, Figure 1). The site is approximately 150–270 m above sea level and lies on a north slope with lateritic soil (pH 4.0). The study area has a south subtropical monsoon climate with an average annual temperature of 21.2°C and mean annual rainfall of 1600 mm. The mean annual air humidity is approximately 82%. Five planting densities (2500 ha-1 [2 × 2 m spacing], 1667 ha-1 [2 × 3 m], 1111 ha-1 [3 × 3 m], 833 ha-1 [3 × 4 m], 625 ha-1 [4 × 4 m]) were arranged in a randomized complete block design with four replicates in August 2010. The afforestation costs for five planting densities are shown in Table 1. Each plot was about 0.05 hectare. All trees in the experimental site were from the same provenance. One or two rows of trees of M. laosensis were planted around each plot for protection, so subject trees at plot edges were not influenced by neighboring plot circumstances.
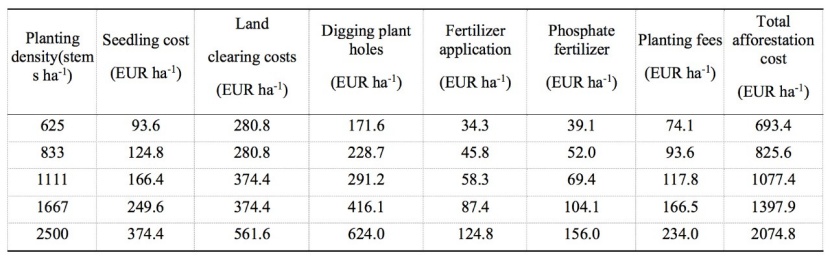
Table 1: Plantation costs for five planting density treatments. / Tabelle 1: Plantagenkosten für fünf Pflanzdichtevarianten.
Measurements
Between July 13 and September 17, 2018 we measured diameter at breast height, tree height, height of the first live branch, and height of the first dead branch. Branch quantity (number, proportion), morphology (diameter, length, and angle), position (height and azimuth), and status (dead or alive) under 12 m of tree height were investigated in 60 dominant or co-dominant trees at five planting densities. Four plots were established for each of the five planting densities (in total 20 plots). Three dominant or co-dominant trees per plot were sampled for branch measurement. All sample trees were surrounded by healthy neighboring trees in all directions and only had a single leader; the trees were disease-free and had no broken tops. For each branch along the stem of the sample trees, the following variables were measured: branch diameter (30 mm from the base of each branch), branch length (length of primary branches), branch angle (from the branch base to the stem; a value of 0° indicates a perpendicular branch), relative branch height, and branch azimuth (0–360°). The azimuth was divided into eight intervals (0–45°, 46–90°, 91–135°,136–180°, 181–225°, 226–270°, 271–315°, and 316–360°). The status of each branch (dead or alive) was recorded and the numbers of live and dead branches were counted from the ground to the top of the sample tree. Live branches were defined as branches with green leaves; dead branches were defined as branches with no green leaves.
Researchers used an extendable ladder to measure all branches on each tree in situ. The branch height and branch length were measured with a steel tape after using chalk to mark the stem at every 1 m, branch diameter was measured with an electronic Vernier caliper, and branch angle was measured with an electronic protractor. In total, 4363 branches (3312 dead branches and 1051 live branches) were measured from 60 dominant or co-dominant trees (3 trees per plot × 4 plots per treatment × 5 treatments).
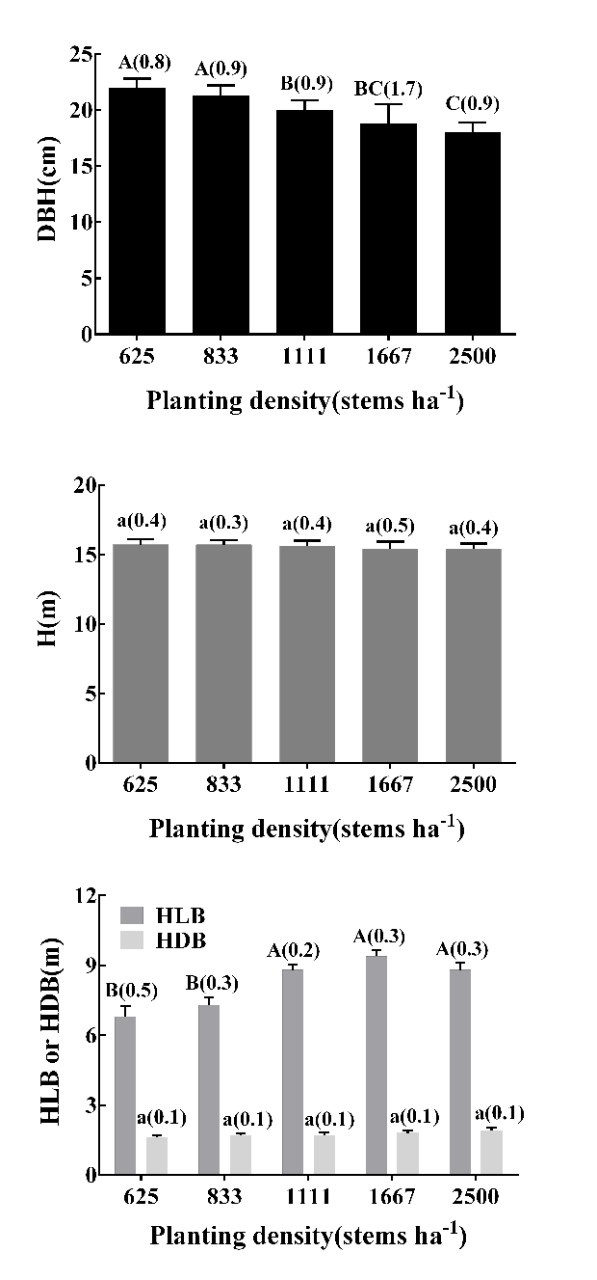
Figure 2: Diameter at breast height (DBH), tree height (H), height to first living branch (HLB) and height to dead branch (HDB) and associated analysis of variance of planting densities on dominant and co-dominant tree in a 8-year-old M. laosensis plantation. Standard error is shown in parentheses; different letters in the same column indicate significant differences between planting density treatments at the 0.01 level. / Abbildung 2: Brusthöhendurchmesser (DBH), Baumhöhe (H), Höhe des ersten lebenden Astes (HLB) und Höhe des ersten toten Astes (HDB) und deren Varianzanalyse bei unterschiedlichen Pflanzdichten von dominanten und kodominanten Bäumen einer 8-jährigen M. laosensis Plantage. Standardfehler in Klammer, unterschiedliche Buchstaben zeigen signifikante Unterschiede zwischen den Pflanzdichten bei 0.01 Signifikanzniveau.
Data Analysis
One-way analysis of variance (ANOVA) and Duncan’s multiple range tests were performed to explore the differences in branch diameter, branch length, branch angle, number of live branches, and number of dead branches on stems below 12 m height among the five planting density treatments. The height of 12 m was used as the timber portion between the base of the stem up to a height of 6–8 m constitutes 90% of the future trees of commercial value (Kint et al. 2010). We used SPSS 20.0 (SPSS, Inc., Chicago, IL, USA) to determine the differences in height of first live branch, height of first dead branch, number of dead branches, number of live branches, number of total branches, and share of dead branches (ratio of dead branch quantity to total branches) among different treatments. We then analysed differences in branch diameter, branch length, branch angle, and branch numbers at different azimuth and heights by planting density.
Results
Tree growth and height of branches
Planting density significantly affected the stem diameter at breast height as well as height of first living branch for dominant or co-dominant trees in the M. laosensis plantation (Fig. 2). The stem diameter at breast height was negatively correlated with planting density from 22.0 to 18.0 cm, and stem diameter at breast height of 625 and 833 stems ha-1 treatment was significantly higher than those of other treatments (P < 0.01). The height of first living branch increased with the increasing planting density from 6.8 to 9.4 m. The treatment of 1667 stems ha-1 was not significantly (6.8%) higher than that of 2500 stems ha-1(P > 0.05). Thus, our results indicated that planting density did not significantly affect the height of first dead branch and tree height of dominant or co-dominant trees.
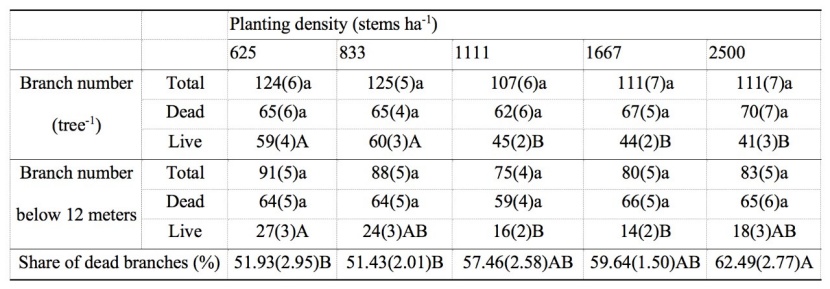
Table 2: The main effect from analysis of variance of planting densities on branch number and share of dead branches of sample dominant or co-dominant trees of M. laosensis plantation. For details, we refer to table 1. / Tabelle 2: Der Haupteffekt der Varianzanalyse der Pflanzdichten auf die Anzahl der Äste und des Anteils abgestorbener Äste von dominanten und kodominanten Bäumen der M. laosensis Plantage. Für Details verweisen wir auf Tabelle 1.
Branch quantity
Planting density had no significant effect on the number of total branches, number of dead branches, and number of dead branches under 12 m among the five density treatments (P > 0.05). The number of live branches and number of live branches below 12 m were negatively correlated with planting density. The share of dead branches was positively correlated with increasing planting density. The highest density of dead branches was observed in the 2500 stems ha-1 treatment (Table 2), whereas there were no significant differences between the 1111, 1667, and 2500 stems ha-1 treatments.
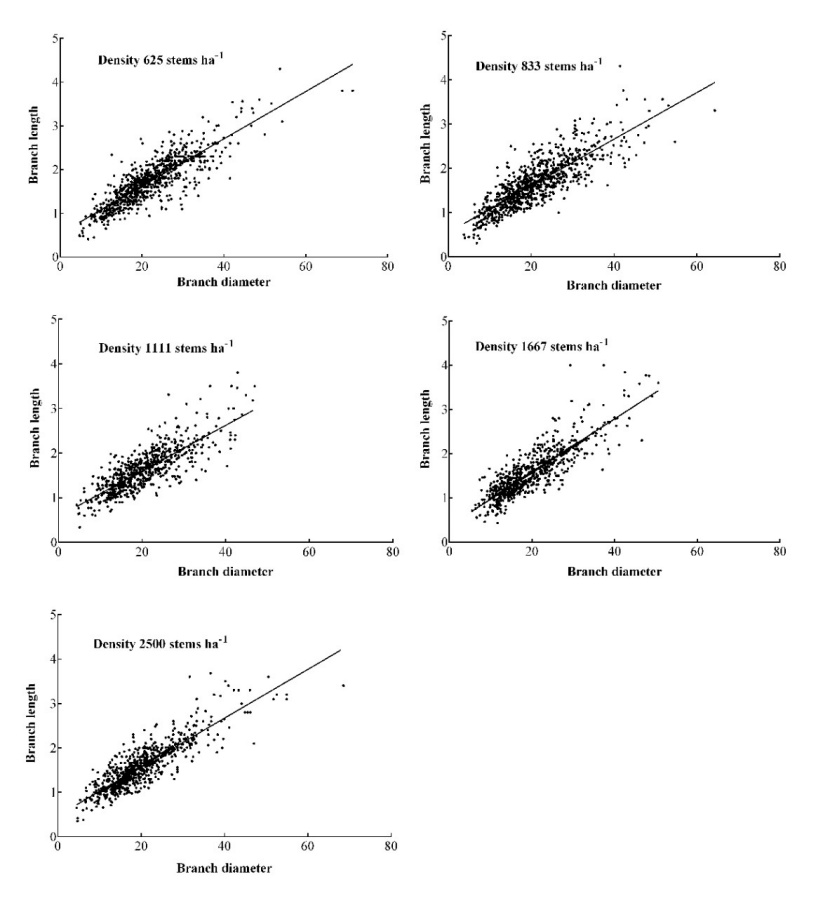
Figure 3: The positive correlation between branch diameter and branch length. / Abbildung 3: Die positive Korrelation zwischen Astdurchmesser und Astlänge.
Branch diameter, length, and angle
Planting density significantly affected the mean branch diameter and branch length of dead branches under 12 m of the stem for M. laosensis (P < 0.01). The mean branch diameter and branch length ranged from 17.95 to 19.32 mm and 1.47 to 1.61 m, respectively, and both showed a decrease with increased planting density. There were no significant differences in the mean dead branch diameter and branch length between 1667 and 2500 stems ha-1 treatments, and the branch diameter and branch length of the 1667 stems ha-1 treatment were only 2.1% and 1.4%, respectively, which were greater than those of the 2500 stems ha-1 treatment. However, the mean branch diameter and branch length of living branches were not significantly affected by planting density. In addition, there was a significant linear positive relationship between branch diameter and branch length (Fig. 3).
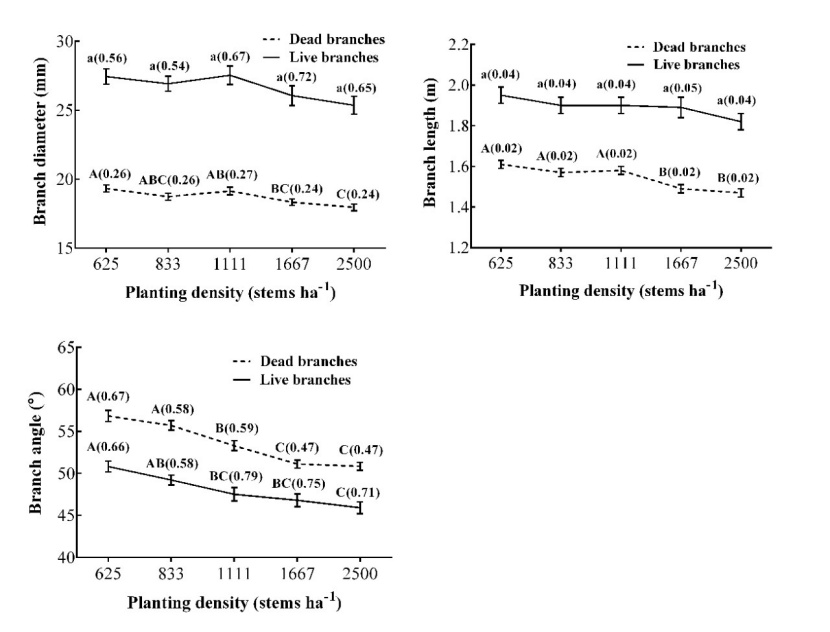
Figure 4: The main effect from analysis of variance of planting densities on branch diameter, length, and angle of dominant or co-dominant trees in the M. laosensis plantation. For details, we refer to table 1. / Abbildung 4: Der Haupteffekt aus der Varianzanalyse der Pflanzdichten auf Durchmesser, Länge und Winkel der Äste von dominanten oder kodominanten Bäume in der M. laosensis Plantage. Für Details verweisen wir auf Tabelle 1.
The mean branch angles of both dead and living branches under 12 m were negatively correlated with planting density. The angles ranged from 50.85° to 56.84° and 45.91° to 50.82°, respectively. The branch angle at the lower density (625 and 833 stems ha-1) was significantly larger than that in the other treatments, indicating that competition limits horizontal space available for branch growth with increasing planting density. The branch angle of living branches was more acute than that of dead branches at the same planting density (Fig. 4).
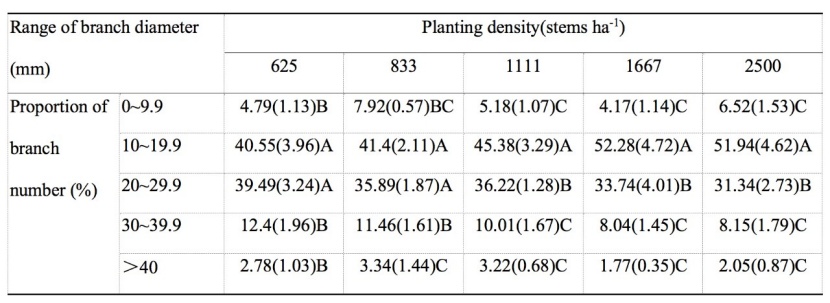
Table 3: The main effect from analysis of variance of planting densities on proportion of branch number for different ranges of branch diameters of dominant or co-dominant trees in the M. laosensis plantation. For details, we refer to table 1. / Tabelle 3: Der Haupteffekt der Varianzanalyse der Pflanzdichte auf den Astanteil für verschiedene Astdurchmesserbereiche von dominanten oder kodominanten Bäume in der M. laosensis Plantage. Für Details verweisen wir auf Tabelle 1.
The proportions of branch number on the lower stem (≤12 m height) of the M. laosensis plantation first increased and then decreased with an increasing range of branch diameter at the same planting density. The branch number at the same planting density decreased in the order 10–19.9 > 20–29.9 > 30–39.9 > 0–9.9 mm. The proportion of branches with a branch diameter greater than 40 mm was lowest, showing a value of only 1–4% among the five planting density treatments. The proportion of branch number in the range of 10–19.9 mm was significantly different from that of 20–29.9 mm at medium and high densities (1111, 1667, and 2500 stems ha-1, P < 0.01). We noted no significant difference at low densities (625 and 833 stems ha-1), indicating that the frequency of the number of larger branches increased with decreasing planting density (Table 3). In contrast, the number of branches with diameters of 20–29.9 mm and 30–39.9 mm tended to decrease with increasing planting density. However, the number of branches with a diameter of 10–19.9 mm increased with increasing planting density and reached a maximum in the 1667 stems ha-1 planting density treatment.
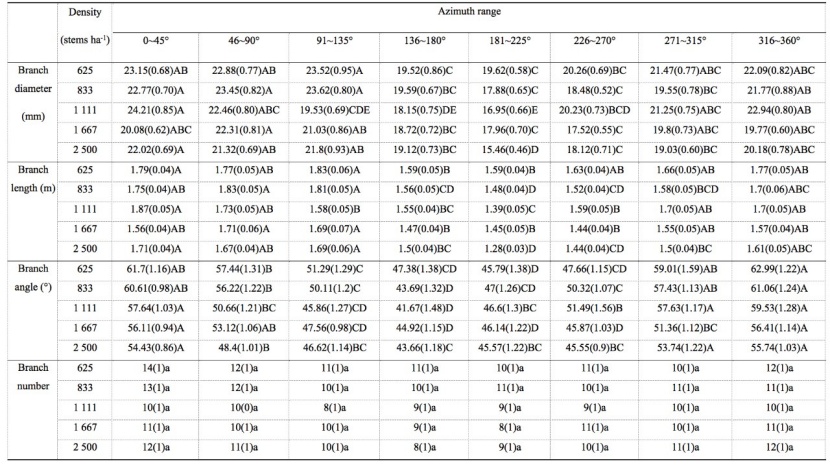
Table 4: The main effect from analysis of variance of different azimuth on branch characteristics for M. laosensis under the same planting density. For details, we refer to table 1. / Tabelle 4: Der Haupteffekt aus der Varianzanalyse verschiedener Azimutwinkel auf die Asteigenschaften von M. laosensis unter der selben Pflanzdichte. Für Details verweisen wir auf Tabelle 1.
Branch Distribution
By analyzing the distribution of branch diameter, branch length, branch angle, and branch quantity with a different azimuth at the same planting density, we found that branches pointing towards the north had a larger diameter, longer branches, higher branch angle, and a higher number of branches than those pointing towards the south. The branch diameter and branch length of the five planting density treatments first decreased and then increased with an increasing azimuth angle. They reached a maximum value in the interval of 0–45° or 46–90° with good light conditions and decreased to a minimum value in the interval of 136–180° or 181–225°. Variance analysis showed no significant difference between the branch diameter and branch length in azimuth intervals 0–45°, 46–90°, 91–135°, and 316–360°. The branch angle reached a maximum value in the interval of 0–45° or 316–360° and decreased to a minimum value in the interval of 136–180°. The influence of different azimuth on branch number was not significant for the five planting density treatments (Table 4). Although branch number was not significantly affected by planting density, this value first decreased and then increased with increasing azimuth.
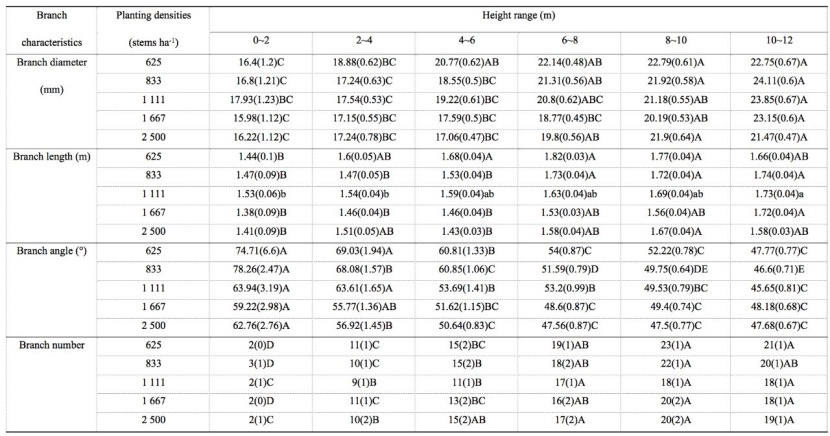
Table 5: The main effect from analysis of variance of different height on branch characteristics for M. laosensis under the same planting density. For details, we refer to table 1. / Tabelle 5: Der Haupteffekt aus der Varianzanalyse unterschiedlicher Höhe auf Asteigenschaften für M. laosensis unter der gleichen Pflanzdichte. Für Details verweisen wir auf Tabelle 1.
The branch diameter and branch length under the same planting density increased with increasing relative branch height, after which there was no significant change. The branch diameter reached a maximum at 8–10 or 10–12 m and decreased to a minimum at 0–2 or 2–4 m. The extreme difference in the vertical direction of branch diameter between the five planting density treatments was 5.68–7.31 mm. The branch length reached a maximum at 6–8, 8–10, or 10–12 m and decreased to a minimum at 0–2 or 2–4 m. The variation amplitude of branch length at each density in the vertical direction was 0.16–0.38 m. The branch angle and branch number for the same density treatment increased with increasing relative branch height (Table 5).
Discussion
Height of first branch
Planting density had a significant effect on the stem diameter at breast height of the sampled dominant or co-dominant trees. Our results indicate that tree growth in the horizontal direction was inhibited by competition. Tree height was not significantly affected by planting density. This result is consistent with those of studies of planting or initial density and spacing for tree species such as Eucalyptus pilularis and E. cloeziana (Alcorn et al. 2007), Betula alnoides (Wang et al. 2018), six-year-old M. laosensis (Zhang et al. 2018), and young Pseudotsuga menziesii (Mirbel) Franco (Li et al. 2007). However, analysis of Pinus taeda L. by Antón-Fernández (2011) revealed differences between the dominant height and spacing, and these differences did not disappear with age. This may be because of the different characteristics of the tree species. Zhang (2018) found that dominant height was significantly affected in the first six years of M. laosensis growth.
There was no significant relation between planting densities and height of first dead branch, whereas height of first living branch was significantly increased with increasing planting density. This confirmed the results of Sun’s study (2014) of Cunninghamia lanceolata, Mäkinen’s study (1999b) of Pinus sylvestris, and Alcorn’s study (2007) on E. pilularis and E. cloeziana, demonstrating that the mortality of branches on the lower stem increased with increasing stand density. However, the self-pruning ability of M. laosensis was lower than that of other tree species. Most dead branches on the lower stem were dry but not easily detached from the stem. Therefore, the capacity for self-pruning cannot be determined by the single indicator of the height of the first living branch but should also be comprehensively evaluated with multiple indicators, such as the height of the first dead branch and numbers of dead branches, living branches, and total branches.
Branch quantity
Branch quantity, particularly the number of dead branches hanging on the stem was closely related to wood quality. The number of total branches and number of dead branches in the 8-year old M. laosensis plantation did not differ significantly between the five planting density treatments. This result agreed with those of previous studies on planting density for some broad-leaf tree species such as 8-year old and 14-year-old B. alnoides (Wang et al. 2016b, 2018) and Fagus sylvatica (Kint et al. 2010). Our previous study of standard trees of an 8-year old M. laosensis plantation, Mäkinen and Hein’s study (2006) of Picea abies (L.) Karst, and Hein’s study (2008) of Pseudotsuga menziesii also showed that the branch number was mainly controlled by genes and a low correlation with environmental factors. Therefore, cultivating non-knot or lower-knot timber of M. laosensis by using density to control the branch number cannot be considered.
The number of live branches decreased significantly with increasing planting density. This result agreed with those of studies of E. pilularis and E. cloeziana (Alcorn et al. 2007), and B. alnoides (Wang et al. 2018), indicating that the forest environment affected the preservation of living branches. Light conditions at low density are better than those in higher density forests. Branches in the lower density groups have a wider growth space and longer survival time.
Branch characteristics
The branch diameter and branch length of dead branches were directly related to the formation of dead-knots. The branch diameter and branch length of dead branches of M. laosensis on the lower stem (≤12 m height) decreased with increasing planting densities, whereas those of living branches were not significantly affected by planting densities. This result is consistent with those of studies of 10-year-old B. alnoides (Wang et al. 2016a), 20-year-old Pinus sylvestris (Gort et al. 2010), young Pseudotsuga menziesii (Mirbel) Franco (Newton et al. 2012), and 4-year-old Eucalyptus robusta (Alcorn et al. 2007). Our previous study of standard trees of M. laosensis also showed that branch diameter and branch length were negatively correlated with planting density. However, the difference in branch diameter and branch length of standard trees at different planting densities was mainly reflected in the lower density (625, 833 stems ha-1), medium density (1111, 1667 stems ha-1), and higher density (2500 stems ha-1), whereas the difference in dominant or co-dominant trees was reflected at lower density (625, 833 and 1111 stems ha-1) and higher density (1667 and 2500 stems ha-1). Because branch size was significantly positively correlated with the size of dead-knots in the timber, afforestation at high density may inhibit increases in branch diameter and reduce dead-knot wood problems in M. laosensis. Low density was conducive to cultivation of large-diameter timber, whereas higher density inhibited increases in branch size. The differences in branch diameter and branch length between the two higher planting density treatments (1667 and 2500 stems ha-1) was not significant, whereas the afforestation cost of 1667 stems ha-1 density treatment was 32.62% lower than that at the 2500 stems ha-1 density.
The branch angle of both dead branches and living branches significantly increased with decreasing planting density. This may be because the reduction in planting density resulted in a larger space for growth and lowered the competition among branches in the horizontal direction. For M. laosensis, larger branches were heavier than smaller branches because of their larger number of leaves. Hence, the vertical pull of gravity may increase branch angle. Branch diameter was negatively related to planting density. Branches in the low-density forest were subjected to gravity in the vertical direction, which can also increase the angle between the branches and stem. In contrast, a larger branch angle made it easier for leaves to absorb sunlight. This result partly agrees with our study of standard trees of M. laosensis and Alcorn et al. (2007) and Henskens et al. (2001) studies of some eucalyptus species. However, Gort et al. (2010) evaluated Pinus sylvestris and found that branch angle was controlled by genes and not significantly affected by density. Wang et al. (2018) found in B. alnoides that although the branch angle was not significantly affected by planting density, lower planting density increased the branch angle.
Branch Distribution
In the horizontal direction, the branch diameter, branch length, and branch angle of M. laosensis on the side exposed to higher levels of sun at the same planting density and different azimuth were larger than those in the shade. The branch number was not significantly affected by the different azimuth range. Wang et al. (2018) evaluated 5-year-old B. alnoides and Xiao et al. (2006) evaluated Pinus sylvestris L. var. mongolica Litv. and found that the azimuth had no discernible effect on the number of primary branches. This result differs from our previous study of standard trees of an 6-year old M. laosensis plantation, in which the branch number of standard trees first decreased and then increased significantly in the horizontal direction with increasing azimuth. The dominant or co-dominant trees were taller than the rest, enabling the capture of more sunlight. For the large trees, the distribution of branches around the trunk was affected by the prevailing light conditions but not significantly. These branch indicators generally reached a maximum value in the azimuth angle of 0–45° but the maximum value of some densities were observed in the azimuth of 316–360° interval. This is because the slope of the test location is not an absolute north slope; there were subtle changes in the slope, which slowly changed the northeast slope into a northwest slope.
In the vertical direction, the branch number first increased gradually and then stabilized with increasing relative branch height. This result is consistent with that of Liu and Li (2007) for Pinus sylvestris L. var. mongolica Litv., where branches on the lower stems withered more than those in the crown. The branch diameter and branch length were significantly affected by the relative branch height. This finding partly agrees with those of studies of Betula alnoides (Wang et al. 2016b) and E. pilularis and E. grandis (Kearney et al. 2007), in which branch diameter and branch length increased significantly and branch angle decreased with increasing relative branch height. This also agrees with our previous study of standard trees of M. laosensis. For Pseudotsuga menziesii (Mirbel) Franco, Weiskittel et al. (2007) found that the branch angle was significantly negatively correlated with branch height.
Economic implications
Our results suggest that planting density should be determined according to the purpose of forest management or forest density should be adjusted by intermediate felling to control the branch size. Artificial pruning should be performed in young M. laosensis plantation when the branch diameter is low and the branches are rapidly growing. At this stage, the wound heals quickly and is less likely to be infected after pruning. Afforestation at a density of 1667 stems ha-1 can produce only approximately 2.1% larger branches than at 2500 stems ha-1, but 32.62% of the cost could be saved on afforestation compared to that at 2500 stems ha-1. Our findings provide a reference for density control, intensive management, and artificial pruning of M. laosensis plantations to enable production of high-quality and large-diameter timber. However, this was a static study of branch development in an 8-year-old M. laosensis plantation. As the fundamental aim of our study was to cultivate non-knot timber and improve timber quality, further studies of the relationship between phenotype features and knot formation are necessary. Additionally, the dynamic growth process of branch development with age should be evaluated.
Acknowledgements
The authors would like to thank Chun-sheng Wang for participating in the design of the experiment and Dr. Mathias Neumann for providing valuable advice on this paper. This work was financially supported by the Fundamental Research Funds for the Central Non-profit Research Institutes of CAF (CAFYBB2019SZ002) and Forestry Science and Technology Innovation Project of Guangdong Province (2019KJCX004).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
Alcorn PJ, Pyttel P, Bauhus J, Smith RG, Thomas D, James R, Nicotra A (2007) Effects of initial planting density on branch development in 4-year-old plantation grown Eucalyptus pilularis and Eucalyptus cloeziana trees. For Ecol Manag 252:41-51
Antón-Fernández C, Burkhart HE, Strub M, Amateis RL (2011) Effects of initial spacing on height development of loblolly pine. For Sci 57:201-211
Biging GS, Dobbertin M (1992) A comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees. Forestence 38:695-720
Chen L, Zeng J, Jia HY, Zeng J, Guo WF, Cai DX (2012) Growth and nutrient uptake dynamics of Mytilaria laosensis seedlings under exponential and conventional fertilization. Soil Sci Plant Nutr 58:618-626
David C. Mercker. (2007). Technical guide to crop tree release in hardwood forests. Professional hradwood notes 1-24
De Freitas, Joberto Veloso (2004). Improving tree selection for felling and retention in natural forest in amazonia through spatial control and targeted seed tree retention:a case study of a forest management project in amazonas state, brazil. University of Aberdeen
Gort J, Zubizarreta-Gerendiain A, Peltola H, Kilpeläinen A, Pulkkinen P, Jaatinen R, Kellomäki S (2010) Differences in branch characteristics of Scots pine (Pinus sylvestris L.) genetic entries grown at different spacing. Ann For Sci 67:705
Guo WF, Cai DX, Jia HY, LI YX, LU ZF (2006) Growth laws of Mytilaria laosensis plantation. For Res 19:585-589
Hein S (2008) Knot attributes and occlusion of naturally pruned branches of Fagus sylvatica. For Ecol Manag 256:2046-2057
Hein S, Spiecker H (2007) Comparative analysis of occluded branch characteristics for Fraxinus excelsior and Acer pseudoplatanus with natural and artificial pruning. Can J For Res 37:1414-1426
Hein S, Weiskittel AR, Kohnle U (2008) Effect of wide spacing on tree growth, branch and sapwood properties of young Douglas-fir [Pseudotsuga menziesii (Mirb.) Franco] in south-western Germany. Eur J For Res 127:481-493
Henskens FL, Battaglia M, Cherry ML, Beadle CL (2001) Physiological basis of spacing effects on tree growth and form in Eucalyptus globulus. Trees 15:365-377
Johansson K (1992) Effects of initial spacing on the stem and branch properties and graded quality of Picea abies (L.) karst. Scand J For Res 7:503-514
Kearney D, James R, Montagu K, Smith RG (2007) The effect of initial planting density on branching characteristics of Eucalyptus pilularis and E. grandis. Aust For 70:262-268
Kellomäki S, Oker-Blom P, Valtonen E, Väisänen H (1989) Structural development of Scots pine stands with varying initial density: Effect of pruning on branchness of wood. For Ecol Manag 27:219-233
Kint V, Hein S, Campioli M, Muys B (2010) Modelling self-pruning and branch attributes for young Quercus robur L. and Fagus sylvatica L. trees. For Ecol Manag 260:2023-2034
Li Y, Turnblom EC, Briggs DG (2007) Effects of density control and fertilization on growth and yield of young Douglas-fir plantations in the Pacific Northwest. Can J For Res 37:449-461
Liao SX, Li k, Lu YC, Li ZY, Yu JS (2009). Analysis on growth of the dominant population of Pinus yunnanensis in central yunnan plateau 1:80-84
Lin DX, Han JF, Xiao ZQ, Hong CF (2000) Mytilaria laosensis improvement of soil physical and chemical properties. J Fujian College For 22:62-65 (in Chinese)
Liu ZG, Li FR (2007) Modeling of spatial distribution of primary branches within the crowns of Pinus sylvestris stands. Scientia Silvae Sinicae 43:19-27
Mäkinen H (1999a) Effect of stand density on radial growth of branches of Scots pine in southern and central Finland. Can J For Res 29:1216-1224
Mäkinen H (1999b) Growth, suppression, death, and self-pruning of branches of Scots pine in southern and central Finland. Can J For Res 29:585-594
Mäkinen H (2002) Effect of stand density on the branch development of silver birch (Betula pendula Roth) in central Finland. Trees 16:346-353
Mäkinen H, Hein S (2006) Effect of wide spacing on increment and branch properties of young Norway spruce. Eur J For Res 125:239-248
Mäkinen H, Saranpää P, Linder S (2001) Effect of nutrient optimization on branch characteristics in Picea abies (L.) karst. Scand J For Res 16:354-362
Neilsen WA, Gerrand AM (1999) Growth and branching habit of Eucalyptus nitens at different spacing and the effect on final crop selection. For Ecol Manag 123:217-229
Newton M, Lachenbruch B, Robbins JM, Cole EC (2012) Branch diameter and longevity linked to plantation spacing and rectangularity in young Douglas-fir. For Ecol Manag 66:75-82
O'Hara KL (1991) A biological justification for pruning in coastal Douglas-fir stands. Western J Appl For 6:59-63
Sun H, Zhang J, Duan A (2014) Study on the self-pruning of Chinese fir plantation. For Res 27:626-630
Wang CS, Hein S, Zhao ZG, Guo JJ, Zeng J (2016a) Branch occlusion and discoloration of Betula alnoides, under artificial and natural pruning. For Ecol Manag 375:200-210
Wang CS, Tang C, Hein S, Guo JJ, Zhao ZG, Zeng J (2018) Branch development of five-year-old Betula alnoides plantations in response to planting density. Forests 9:42
Wang CS, Zeng J, Hein S, Zhao ZG, Guo JJ, Zeng J (2016b) Crown and branch attributes of mid-aged Betula alnoides plantations in response to planting density. Scand J For Res 32:679-687
Wang XQ, Liu XE, Ren HQ (2007) Effect of crown attributes on wood characteristics and increments of Populus × xiaohei. For Res 20:801-806
Wang ZH, Yin GT, Yang JC, Qin GM (2019) Effects of planting density on branch development of Mytilaria laosensis plantations. For Res 32:78-86
Weiskittel AR, Maguire DA, Monserud RA (2007) Modeling crown structural responses to competing vegetation control, thinning, fertilization, and Swiss needle cast in coastal Douglas-fir of the Pacific Northwest, USA. For Ecol Manag 245:96-109
Xiao R, Li FR, Liu ZG (2006) Branching structure analysis of mongolian pine plantation. Bull Bot Res 26:490-496
Zhang YF, Yin GT, Yang JC, Li RS, Zou WT, Wang X (2018) Effect of planting density on the early growth of Mytilaria laosensis plantation. For Res 31:83-89



